Тим за киропрактику и функционалну медицину клинике за оксидативни стрес. Оксидативни стрес се дефинише као поремећај у равнотежи између производње реактивног кисеоника (слободних радикала) и антиоксидативне одбране. Другим речима, то је неравнотежа између производње слободних радикала и способности тела да се супротстави или детоксификује штетним ефектима кроз неутрализацију антиоксидансима. Оксидативни стрес доводи до многих патофизиолошких стања у телу. То укључује неуродегенеративне болести, тј. Паркинсонову болест, Алцхајмерову болест, мутације гена, карциноме, синдром хроничног умора, крхки Кс синдром, поремећаје срца и крвних судова, атеросклерозу, срчану инсуфицијенцију, срчани удар и инфламаторне болести. Оксидација се дешава под неколико околности:
ћелије користе глукозу за стварање енергије
имуни систем се бори против бактерија и ствара упалу
тела детоксинишу загађиваче, пестициде и цигаретни дим
У нашем се телу у било ком тренутку одвијају милиони процеса који могу довести до оксидације. Ево неколико симптома:
Умор
Губитак памћења и / или мождана магла
Болови у мишићима и зглобовима
Боре заједно са сивом косом
Смањен вид
Главобоља и осетљивост на буку
Сусцептибилност за инфекције
Одабир органске хране и избегавање токсина у вашем окружењу чини велику разлику. Ово, заједно са смањењем стреса, може бити корисно у смањењу оксидације.
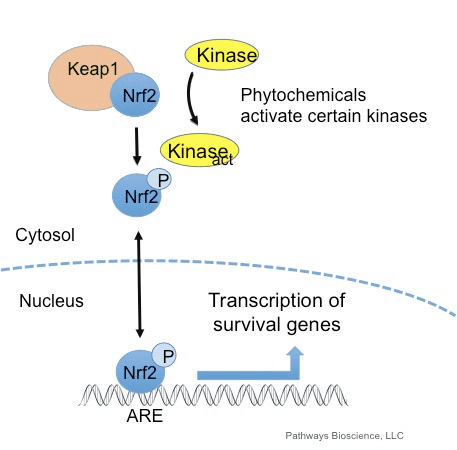
Оксиданти се углавном производе на контролисан начин како би се регулисали основни процеси у људском телу, укључујући поделу ћелија, упале, имунолошку функцију, аутофагију и одговор на стрес. Међутим, неконтролисана производња ових оксиданата може томе допринети оксидативни стрес, што може утицати на ћелијску функцију, што доводи до развоја токсичности, хроничне болести и рака. Заштитни антиоксидативни механизми људског тела су регулисани низом виталних путева који контролишу одговор ћелије на оксидансе. Фактор везан за нуклеарни фактор еритроид 2, иначе познат као Нрф2, је нови регулатор ћелијске отпорности на оксидансе. Сврха чланка у наставку је да се дискутује и демонстрира нова улога Нрф2 у функцији митохондрија.
Апстрактан
Фактор 2 транскрипције НФ-Е45 п2 везан за фактор 2 (Нрф2; назив гена НФЕ2Л2) омогућава адаптацију и преживљавање у условима стреса регулацијом експресије гена различитих мрежа цитопротективних протеина, укључујући антиоксидативне, антиинфламаторне и ензиме за детоксикацију. као протеини који помажу у поправљању или уклањању оштећених макромолекула. Нрф2 има кључну улогу у одржавању ћелијске редокс хомеостазе регулацијом биосинтезе, коришћења и регенерације глутатиона, тиоредоксина и НАДПХ и контролом производње реактивних врста кисеоника од стране митохондрија и НАДПХ оксидазе. У хомеостатским условима, Нрф2 утиче на потенцијал митохондријалне мембране, оксидацију масних киселина, доступност супстрата (НАДХ и ФАДХ2/сукцинат) за дисање и синтезу АТП-а. У условима стреса или стимулације фактора раста, активација Нрф3 се супротставља повећаном стварању реактивних врста кисеоника у митохондријама преко транскрипционе регулације одспојног протеина 1 и утиче на биогенезу митохондрија одржавањем нивоа нуклеарног респираторног фактора 1 и рецептора активираног пролифератором пероксизома? коактиватор 2?, као и промовисањем биосинтезе пуринских нуклеотида. Фармаколошки Нрф1,4 активатори, као што је природни изотиоцијанат сулфорафан, инхибирају оксиданс посредовано отварање прелазних пора митохондријалне пермеабилности и отицање митохондрија. Занимљиво је да је синтетичко 1,2,3-дифенил-2-триазолно једињење, првобитно дизајнирано као активатор Нрф2, пронађено да промовише митофагију, чиме доприноси укупној митохондријалној хомеостази. Дакле, НрфXNUMX је истакнут играч у подршци структурном и функционалном интегритету митохондрија, а ова улога је посебно кључна у условима стреса.
Нрф2 има пресудну улогу у одржавању ћелијске редокс хомеостазе.
Нрф2 утиче на потенцијал митохондријске мембране и синтезу АТП.
Нрф2 утиче на оксидацију митохондријских масних киселина.
Нрф2 подржава структурни и функционални интегритет митохондрија.
Активатори Нрф2 имају благотворне ефекте када је митохондријска функција угрожена.
увод
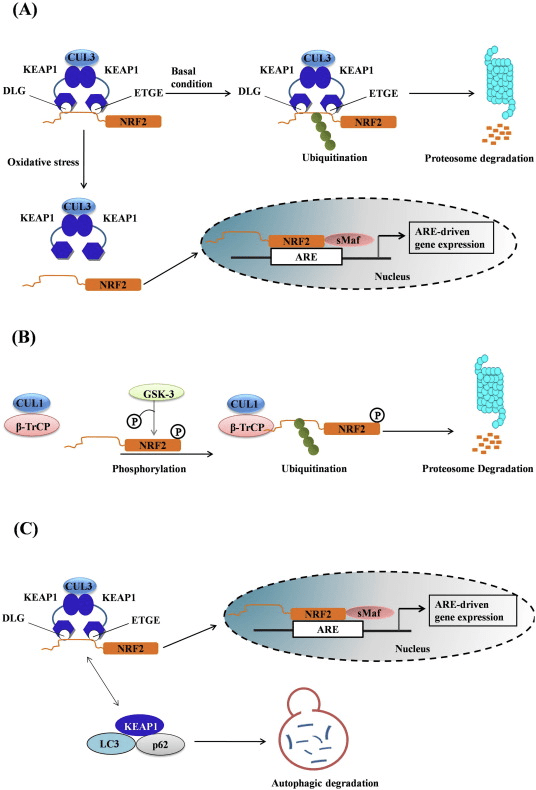
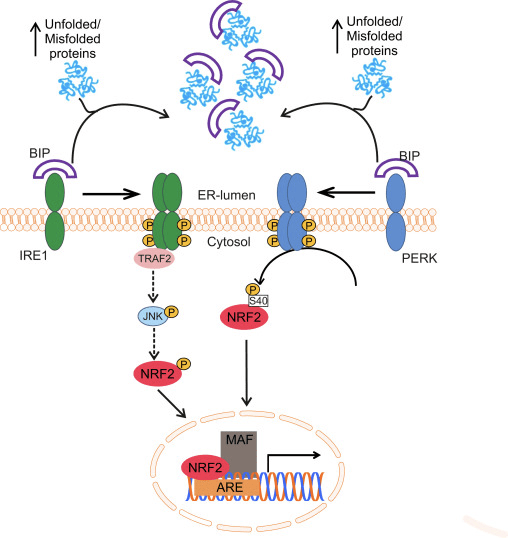
Фактор транскрипције НФ-Е2 п45 повезан са фактором 2 (Нрф2; назив гена НФЕ2Л2) регулише експресију мрежа гена који кодирају протеине са различитим цитопротективним активностима. Сам Нрф2 се контролише првенствено на нивоу стабилности протеина. У базалним условима, Нрф2 је краткотрајни протеин који је подвргнут континуираној убиквитинацији и протеасомској деградацији. Постоје три позната система убиквитин лигазе који доприносе разградњи Нрф2. Историјски гледано, први негативни регулатор Нрф2 који је откривен био је Келцх-сличан ЕЦХ-повезани протеин 1 (Кеап1) [1], протеин супстрата-адаптера за Цуллин 3 (Цул3) / Рбк1 убикуитин лигазу [2], [3], [ 4]. Кеап1 користи високо ефикасан циклички механизам за циљање Нрф2 за убиквитинацију и протеасомску деградацију, током које се Кеап1 континуирано регенерише, омогућавајући циклус да се настави (слика 1А) [5]. Нрф2 је такође подвргнут разградњи посредованој гликоген синтаза киназом (ГСК) 3 / а - ТрЦП-зависна убиквитин лигаза на бази Цул1 [6], [7]. Недавно је забележено да је, током стања стреса ендоплазматског ретикулума, Нрф2 убиквитиниран и разграђен у процесу посредовања од Е3 убиквитин лигазе Хрд1 [8].
Слика 1 Модел цикличног секвенцијалног везивања и регенерације за разградњу Нрф1 посредовану Кеап2. (А) Нрф2 се веже узастопно за бесплатни Кеап1 димер: прво кроз свој домен везивања ЕТГЕ (црвени штапићи) са високим афинитетом, а затим кроз домен везивања ДЛГ (црни штапићи) са малим афинитетом. У овој конформацији протеинског комплекса, Нрф2 се подвргава убиквитинацији и циља на протеасомску разградњу. Бесплатни Кеап1 се регенерише и може да се веже за ново преведени Нрф2 и циклус започиње поново. (Б) Индуктори (бели дијаманти) реагују са сензорским цистеинима Кеап1 (плави штапићи), што доводи до промене конформације и оштећења активности адаптера супстрата. Слободни Кеап1 се не регенерише, а ново синтетизовани Нрф2 се акумулира и транслоцира у језгро.
Поред тога што служи као протеин протеина адаптера убиквитин лигазе, Кеап1 је и сензор за широк спектар малих молекулских активатора Нрф2 (названих индуктори) [9]. Индуктори блокирају циклус разградње Нрф1 посредоване Кеап2 хемијском модификацијом специфичних остатака цистеина унутар Кеап1 [10], [11] или директним ометањем интерфејса везивања Кеап1: Нрф2 [12], [13]. Сходно томе, Нрф2 се не разграђује, а фактор транскрипције се акумулира и транслоцира у језгро (слика 1Б), где формира хетеродимер са малим Маф протеином; везује се за елементе реакције на антиоксидант, горње регулаторне регије својих циљних гена; и покреће транскрипцију [14], [15], [16]. Батерија Нрф2 циљева садржи протеине са различитим цитопротективним функцијама, укључујући ензиме метаболизма ксенобиотика, протеине са антиоксидативним и противупалним функцијама и протеасомске подјединице, као и протеине који регулишу ћелијску редокс хомеостазу и учествују у посредничком метаболизму.
Нрф2: главни регулатор ћелијске редокс хомеостазе
Функција Нрф2 као главног регулатора ћелијске редокс хомеостазе је широко призната. Експресија гена и каталитичке и регулаторне подјединице ?-глутамил цистеин лигазе, ензима који катализује корак који ограничава брзину у биосинтези редукованог глутатиона (ГСХ), директно је регулисана Нрф2 [17]. кЦТ подјединица система кц-, која увози цистин у ћелије, такође је директна мета транскрипције Нрф2 [18]. У ћелији, цистин се претвара у цистеин, прекурсор за биосинтезу ГСХ. Поред своје улоге у биосинтези ГСХ, Нрф2 обезбеђује средства за одржавање глутатиона у редукованом стању путем координисане транскрипционе регулације глутатион редуктазе 1 [19], [20], која редукује оксидовани глутатион у ГСХ коришћењем редукционих еквивалената из НАДПХ. . Потребни НАДПХ обезбеђују четири главна ензима који стварају НАДПХ, јабучни ензим 1 (МЕ1), изоцитрат дехидрогеназа 1 (ИДХ1), глукоза-6-фосфат дехидрогеназа (Г6ПД) и 6-фосфоглуконат дехидрогеназа (ПГД), од којих су сви транскрипционо делимично регулисан Нрф2 (слика 2) [21], [22], [23], [24]. Занимљиво је да Нрф2 такође регулише индуцибилну експресију гена цитосолних, микрозомалних и митохондријалних облика алдехид дехидрогеназе [25], који користе НАД(П)+ као кофактор, што доводи до НАД(П)Х. Заиста, нивои НАДПХ и НАДПХ/НАДП+ омјера су нижи у ембрионалним фибробластима изолованим од Нрф2-нокаут (Нрф2-КО) мишева у поређењу са ћелијама из њихових парњака дивљег типа (ВТ), а нивои НАДПХ опадају након Нрф2 нокаута у ћелијске линије рака са конститутивно активним Нрф2 [26]. Као што се и очекивало, нивои ГСХ су нижи у ћелијама у којима је Нрф2 поремећен; обрнуто, активација Нрф2 генетским или фармаколошким путем доводи до повећања ГСХ [27], [28], [29]. Важно је да Нрф2 такође регулише експресију гена тиоредоксина [30], [31], [32], тиоредоксин редуктазе 1 [28], [29], [32], [33] и сулфиредоксина [34], који су неопходни за редукцију оксидисаних протеинских тиола.
Слика 2 Улога Нрф2 у метаболизму ћелија које се брзо шире. Нрф2 је позитивни регулатор гена који кодирају ензиме и у оксидативном краку [тј. Глукоза-6-фосфат дехидрогеназа (Г6ПД) и у 6-фосфоглуконат дехидрогенази (ПГД)] и у неоксидативном краку [тј., У трансалдолази 1 (ТАЛДО1) и у транскетолази ( ТКТ)] пентозо-фосфатног пута. Г6ПД и ПГД генеришу НАДПХ. Нрф2 такође регулише експресију гена друга два ензима који генеришу НАДПХ, јабучни ензим 1 (МЕ1) и изоцитрат дехидрогеназу 1 (ИДХ1). Експресију гена фосфорибозил пирофосфат амидотрансферазе (ППАТ), која катализује улазак у биосинтетски пут де ново пурина, такође позитивно регулише Нрф2, као и израз метиленететрахидрофолат дехидрогеназе 2 (МТХФД2), митохондријског ензима са критичном улогом у обезбеђујући једнокарбонске јединице за де ново биосинтезу пурина. Нрф2 негативно регулише пируват киназу (ПК) и очекује се да фаворизује накупљање гликолитичких интермедијера и, заједно са Г6ПД, метаболита који пролази каналом кроз пентоза фосфат и синтезу нуклеинских киселина, аминокиселина и фосфолипида. Нрф2 негативно регулише експресију гена АТП-цитрат лиазе (ЦЛ), што може повећати доступност цитрата за употребу митохондрија или (путем изоцитрата) за ИДХ1. Црвена и плава означавају позитивну, односно негативну регулацију. Митохондрија је приказана сивом бојом. Скраћенице метаболита: Г-6-П, глукоза 6-фосфат; Ф-6-П, фруктоза 6-фосфат; Ф-1,6-БП, фруктоза 1,6-бисфосфат; ГА-3-П, глицералдехид 3-фосфат; 3-ПГ, 3-фосфоглицерат; ПЕП, фосфоенолпируват; 6-П-Гл, 6-фосфоглуконолактон; 6-ПГ, 6-фосфоглуконат; Р-5-П, рибулоза 5-фосфат; ПРПП, 5-фосфорибозил-а-1-пирофосфат; ТХФ, тетрахидрофолат; ИМП, инозин монофосфат; АМП, аденозин монофосфат; ГМП, гванозин монофосфат.
С обзиром на пресудну улогу Нрф2 као главног регулатора ћелијске редокс хомеостазе, није изненађујуће да су, у поређењу са ВТ ћелијама, нивои реактивних врста кисеоника (РОС) виши у ћелијама у којима је Нрф2 поремећен (Нрф2-КО). [35]. Ова разлика је посебно упечатљива код изазивања са агенсима који узрокују оксидативни стрес. Штавише, ћелије са недостатком Нрф2 су много осетљивије на токсичност оксиданата различитих врста и не могу се заштитити индукторима Нрф2, који под истим условима пружају ефикасну и дуготрајну заштиту ВТ ћелијама [29], [36] , [37]. Поред укупне ћелијске редокс хомеостазе, Нрф2 је такође критичан за одржавање митохондријске редокс хомеостазе. Дакле, у поређењу са ВТ, укупни митохондријски НАДХ фонд је значајно повећан у Кеап1-КО и драматично смањен у Нрф2-КО ћелијама [35].
Користећи снимање живих ћелија, недавно смо пратили стопе производње РОС у примарним глионеуроналним културама и резовима можданог ткива изолованим од мишева ВТ, Нрф2-КО или Кеап1-кноцкдовн (Кеап1-КД) [38]. Као што се очекивало, стопа производње РОС била је бржа у ћелијама и ткивима Нрф2-КО у поређењу са њиховим ВТ колегама. Међутим, изнели смо неочекивано запажање да, у поређењу са ВТ, ћелије Кеап1-КД такође имају веће стопе производње РОС, иако је величина разлике између ВТ и Кеап1-КД генотипова била мања од оне између ВТ и Нрф2-КО . Затим смо анализирали ниво мРНК НОКС2 и НОКС4, каталитичке подјединице две изоформе НАДПХ оксидазе (НОКС) које су умешане у патологију мозга, и открили смо да је НОКС2 драматично повећан у условима недостатка Нрф2, док је НОКС4 појачано регулисан када Нрф2 је конститутивно активиран, мада у мањој мери. Квантитативно, величина повећања регулације у ћелијама и ткивима мутираних мишева паралелно је одговарајућем повећању производње РОС [38]. Занимљиво је да Нрф2 не само да регулише НАДПХ оксидазу, већ и РОС који производи НАДПХ оксидаза може да активира Нрф2, као што је приказано у плућним епителним ћелијама и кардиомиоцитима [39], [40]. Даље, врло недавна студија показала је да НАДПХ оксидазно зависна активација Нрф2 представља важан ендогени механизам за заштиту од оштећења митохондрија и ћелијске смрти у срцу током хроничног преоптерећења [41].
Поред каталитичке активности НАДПХ оксидазе, митохондријско дисање је још један главни унутарћелијски извор РОС-а. Коришћењем митохондрије специфичне сонде МитоСОКС испитали смо допринос РОС-а митохондријског порекла укупној производњи РОС-а у примарним изолованим глионеуроналним културама. од мишева ВТ, Нрф2-КО или Кеап1-КД [38]. Као што се очекивало, Нрф2-КО ћелије су имале веће стопе производње митохондријског РОС од ВТ. У сагласности са налазима за укупну производњу РОС-а, стопе производње митохондријског РОС-а у Кеап1-КД такође су биле веће у поређењу са ВТ ћелијама. Важно је да је блокирање комплекса И ротеноном изазвало драматичан пораст производње митохондријског РОС-а и у ћелијама ВТ и у Кеап1-КД, али није имало ефекта у ћелијама Нрф2-КО. За разлику од очекиваног повећања производње митохондријског РОС-а у ВТ ћелијама након додавања пирувата (да би се повећала доступност НАДХ, повећао потенцијал митохондријске мембране и нормализовало дисање), производња РОС-а се смањила у Нрф2-КО ћелијама. Заједно, ови налази снажно сугеришу да је, у одсуству Нрф2: (и) активност комплекса И поремећена, (ии) оштећена активност комплекса И је због ограничења супстрата и (иии) оштећена активност комплекса И је један од главних разлога за повећану производњу митохондријског РОС-а, вероватно захваљујући обрнутом протоку електрона из комплекса ИИ.
Нрф2 утиче на потенцијал и дисање митохондријске мембране
Потенцијал митохондријске мембране (?? м) универзални је показатељ здравља митохондрија и метаболичког стања ћелије. У здравој ћелији, м одржава митохондријски респираторни ланац. Занимљиво је да је стабилно изотопско обележавање аминокиселинама у проучавању протеомије засноване на култури у ћелијској линији естрогена људског епитела МЦФ10А на негативни рецептор естрогена показало да је компонента ланца транспортног електрона митохондрија НДУФА4 појачана фармаколошком активацијом Нрф2, док генетска регулација Нрф2 (ноакдауном Кеап1) доводи до смањења регулације подјединица цитокром ц оксидазе ЦОКС2 и ЦОКС4И1 [42]. Студија протеома јетре помоћу дводимензионалне гел електрофорезе и ласерске десорпције / јонизационе масене спектрометрије уз помоћ матрице открила је да Нрф2 регулише експресију подјединице АТП синтазе? [43]. Поред тога, забележено је да митохондријски протеин ДЈ-1, који игра улогу у одржавању активности комплекса И [44], стабилизује Нрф2 [45], [46], иако неуропротективни ефекти фармаколошке или генетске активације од Нрф2 су независни од ДЈ-1 [47]. Међутим, последице ових посматрања на функцију митохондрија нису истражене.
У сагласности са оштећеном активношћу комплекса И у условима недостатка Нрф2, базални ?? м је нижи у ембрионалним фибробластима мишића Нрф2-КО (МЕФ) и култивисаним примарним глионеуроналним ћелијама у поређењу са њиховим ВТ колегама (слика 3, уметак) [35]. Супротно томе, базални ?? м је већи када је Нрф2 генетски конститутивно регулисан (нокаутом или нокаутом Кеап1). Ове разлике у ?? м међу генотиповима указују да на дисање утиче активност Нрф2. Заиста, процена потрошње кисеоника у базалном стању открила је да је, у поређењу са ВТ, потрошња кисеоника мања у Нрф2-КО и Кеап1-КО МЕФ, за ~ 50, односно ~ 35%.
Слика 3 Предложени механизам за угрожену функцију митохондрија у условима недостатка Нрф2. (1) Смањени нивои МЕ1, ИДХ1, Г6ПД и ПГД резултирају нижим нивоима НАДПХ. (2) Нивои ГСХ су такође ниски. (3) Ниска активност МЕ1 може смањити количину пирувата који улази у митохондрије. (4) Генерисање НАДХ је спорије, што доводи до поремећене активности комплекса И и повећане производње РОС у митохондријима. (5) Редукција ФАД у ФАДХ2 у митохондријалним протеинима је такође смањена, смањујући проток електрона од ФАДХ2 до УбК и у комплекс ИИИ. (6) Спорије формирање УбКХ2 може смањити активност ензима сукцинат дехидрогеназе. (7) Повећани нивои РОС могу даље инхибирати активност комплекса ИИ. (8) Мања ефикасност оксидације масних киселина доприноси смањеној доступности супстрата за дисање митохондрија. (9) Гликолиза је појачана као компензациони механизам за смањену производњу АТП-а у оксидативној фосфорилацији. (10) АТП синтаза ради обрнуто да би одржала ??м. Црвена и плава означавају регулацију нагоре и на ниже, респективно. Кутије означавају доступност експерименталних доказа. Уметак приказује слике митохондрија кортикалних астроцита ВТ и Нрф2-КО визуелизоване потенциометријском флуоресцентном сондом тетраметилродамин метил естра (ТМРМ; 25 нМ). Скала трака, 20 м.
Ове разлике у ??м и дисању међу генотиповима се одражавају на стопу искоришћења супстрата за митохондријално дисање. Примена супстрата за циклус трикарбоксилне киселине (ТЦА) (малат/пируват, који заузврат повећавају производњу супстрата комплекса И НАДХ) или метил сукцината, супстрата за комплекс ИИ, изазива постепено повећање ??м у обе ВТ и Кеап1-КД неуроне, али је стопа повећања већа у Кеап1-КД ћелијама. Што је још важније, облици одговора на ове супстрате ТЦА циклуса су различити између два генотипа, при чему је брзо повећање ??м у Кеап1-КД ћелијама након додавања супстрата праћено брзим падом, а не платоом, што сугерише необично брза потрошња подлоге. Ови налази су у блиској сагласности са много нижим (за 50%) нивоима малата, пирувата и сукцината који су примећени након једночасовног пулса [У-70Ц1]глукозе у Кеап13-КО у поређењу са ВТ МЕФ ћелије [6]. У Нрф1-КО неуронима, само пируват је у стању да повећа ??м, док малат и метил сукцинат изазивају благу деполаризацију. Чини се да је ефекат Нрф24 на производњу митохондријалног супстрата главни механизам којим Нрф2 утиче на функцију митохондрија. Митохондријски НАДХ редокс индекс (равнотежа између потрошње НАДХ комплексом И и производње НАДПХ у ТЦА циклусу) је значајно нижи у Нрф2-КО ћелијама у поређењу са њиховим ВТ колегама, а штавише, стопе регенерације базена НАДХ и ФАДХ2 након инхибиције комплекса ИВ (употребом НаЦН) су спорији у мутантним ћелијама.
У митохондријима изолованим од мишјег мозга и јетре, додавање супстрата за комплекс И или за комплекс ИИ јаче повећава брзину потрошње кисеоника када се Нрф2 активира и мање ефикасно када се Нрф2 поремети [35]. Дакле, малат индукује већу стопу потрошње кисеоника у Кеап1-КД у поређењу са ВТ, али је његов ефекат слабији у Нрф2-КО митохондријима. Слично томе, у присуству ротенона (када је инхибиран комплекс И), сукцинат у већој мери активира потрошњу кисеоника у Кеап1-КД у поређењу са ВТ, док је одговор у Нрф2-КО митохондријама смањен. Поред тога, примарне неуронске културе и мишеви Нрф2-КО су осетљивији на токсичност инхибитора комплекса ИИ 3-нитропропионске киселине и малоната, док је интрастријатална трансплантација астроцита који прекомерно експримирају Нрф2 заштитна [48], [49]. Слично томе, Нрф2-КО мишеви су осетљивији на, док генетска или фармаколошка активација Нрф2 има заштитне ефекте против неуротоксичности изазване комплексом И инхибитором 1-метил-4-фенилпиридинијум јона у 1-метил-4-фенил-1,2,3,6, 49-тетрахидропиридински животињски модел Паркинсонове болести [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61], [XNUMX].
Однос респираторне контроле (РЦР), однос стања 3 (стимулисан АДП) и респирације стања 4 (без присуства АДП), смањен је у одсуству Нрф2, али је РЦР сличан између Кеап1-КД и ВТ митохондрија [35 ]. Пошто је РЦР индикација степена спајања активности митохондријалног респираторног ланца на оксидативну фосфорилацију, овај налаз указује да виша стопа дисања у Кеап1-КД митохондријама није последица раздвајања оксидативне фосфорилације. Даље сугерише да је оксидативна фосфорилација ефикаснија када је Нрф2 активиран. Већа стопа дисања у Кеап1-КД митохондријама је у складу са вишим нивоима производње РОС у митохондријима [38] јер веће стопе дисања могу довести до повећаног цурења електрона. Међутим, у условима оксидативног стреса, повећана производња РОС се супротставља Нрф2-зависној транскрипционој регулацији одспојног протеина 3 (УЦП3), која повећава протонску проводљивост унутрашње мембране митохондрија и последично смањује производњу супероксида [62]. Недавно је показано да производ пероксидације липида 4-хидрокси-2-ноненал посредује у Нрф2 зависној регулацији УЦП3 у кардиомиоцитима; ово би могло бити посебно важно за заштиту у условима оксидативног стреса као што су они током исхемијске реперфузије [63].
Нрф2 утиче на ефикасност оксидативне фосфорилације и синтезу АТП-а
У сагласности са ефектом Нрф2 на дисање, у митохондријима мозга и јетре, недостатак Нрф2 резултира смањеном ефикасношћу оксидативне фосфорилације (процењене односом АДП и кисеоника који се троши за синтезу АТП), док активација Нрф2 (Кеап1 -КД) има супротан ефекат [35]. У поређењу са ВТ, нивои АТП су знатно већи у ћелијама са конститутивном регулацијом Нрф2 и нижи када је Нрф2 срушен [64] или поремећен [35]. Даље, употреба инхибитора оксидативне фосфорилације (олигомицин) или гликолизе (јодосирћетна киселина) открила је да Нрф2 мења начин на који ћелије производе АТП. Дакле, у ВТ неуронима олигомицин изазива потпуни пад АТП, а јодооцетна киселина нема даље дејство. Изузетно је то што у Нрф2-КО ћелијама олигомицин повећава ниво АТП-а, који се тада полако, али потпуно, исцрпљује јодооцтеном киселином, што указује да је у одсуству Нрф2 гликолиза, а не оксидативна фосфорилација, главни извор производње АТП-а. Занимљиво је да, упркос повећаној ефикасности оксидативне фосфорилације у ћелијама Кеап1-КД, додавање олигомицина резултира у ~ 80% смањењу нивоа АТП, а јодооцетна киселина изазива даљњих ~ 20% смањења. Дакле, или недостатак Нрф2, или његова конститутивна активација смањује допринос оксидативне фосфорилације и повећава допринос гликолизе синтези АТП. Овај ефекат је посебно изражен када Нрф2 одсуствује и у складу је са зависношћу ?? м од присуства глукозе у медијуму [35] и повишеним нивоима гликолитичких интермедијера (Г-6-П, Ф-6-П , дихидроксиацетон фосфат, пируват и лактат) након обарања Нрф2 [24].
Пораст нивоа АТП након инхибиције Ф1Ф0-АТПазе олигомицином указује да у одсуству Нрф2, Ф1Ф0-АТПаза функционише као АТПаза, а не као АТП синтаза, тј. Делује обрнуто. Такав преокрет у активности највероватније одражава потребу за пумпањем протона преко унутрашње митохондријске мембране у покушају да се одржи ?? м, што је пресудно за функционални интегритет ове органеле. О преокрету функције Ф1Ф0-АТПазе сведочи и примећена деполаризација митохондрија приликом примене олигомицина у ћелије Нрф2-КО, што је у оштрој супротности са хиперполаризацијом која се јавља у њиховим колегама са недостатком ВТ или Кеап1 [35]. Све у свему, чини се да се у условима недостатка Нрф2 АТП производи првенствено у гликолизи, а овај АТП затим делимично користи Ф1Ф0-АТПаза за одржавање ?? м.
Ефекат недостатка Нрф2 на ?? м посебно је изражен када се ћелије инкубирају у медијуму без глукозе, а ум је око 50% нижи у Нрф2-КО у поређењу са ВТ ћелијама [35]. У условима недостатка глукозе, митохондријска оксидација масних киселина (ФАО) је главни добављач супстрата за дисање и оксидативну фосфорилацију, што указује да Нрф2 може утицати на ФАО. Заиста, ефикасност ФАО и за дуголанчану (Ц16: 0) засићену палмитинску киселину масних киселина и за кратководну (Ц6: 0) хексанску киселину већа је у МЕФ Кеап1-КО и изолованих митохондрија срца и јетре него у њиховим ВТ пандани, док је нижи у Нрф2-КО ћелијама и митохондријима [65]. Ови ефекти су такође изузетно релевантни за људе: заиста, забележено је да се метаболичке промене које указују на бољу интеграцију ФАО са активношћу ТЦА циклуса јављају у хуманим интервентним студијама са дијетама богатим глукорафанином, претечом класичног Нрф2 активатора сулфорафана [ 66].
Током првог корака митохондријске ФАО, про-Р водоник а-угљеника одлази као хидрид који смањује ФАД кофактор на ФАДХ2, што заузврат преноси електроне у убикинон (УбК) у респираторном ланцу, што на крају доприноси производњи АТП . Док стимулација ФАО палмитоилкарнитином у одсуству глукозе узрокује очекивано повећање нивоа АТП у ћелијама ВТ и Кеап1-КО, с тим што је раст АТП бржи у ћелијама Кеап1-КО, идентичан третман не доводи до промена АТП у Нрф2-КО МЕФ [65]. Овај експеримент показује да је, у одсуству Нрф2, ФАО потиснут, а поред тога, подразумева сузбијање ФАО као један од разлога за ниже нивое АТП-а у условима недостатка Нрф2 [35], [64].
Пре свега, хумане 293 Т ћелије у којима је Нрф2 ућуткан имају нижу експресију ЦПТ1 и ЦПТ2 [67], две изоформе карнитин палмитоилтрансферазе (ЦПТ), ензима који ограничава брзину у митохондријском ФАО. Слажу се да су нивои мРНК Цпт1 нижи у јетри Нрф2-КО у поређењу са ВТ мишевима [68]. ЦПТ катализује пренос ацил групе дуголанчаног масног ацил-ЦоА из коензима А у л-карнитин и на тај начин омогућава увоз ацилкарнитина из цитоплазме у митохондрије. Иако ово до данас није испитано, могуће је да поред транскрипционих ефеката на експресију ЦПТ1, Нрф2 такође може утицати на функцију овог ензима контролишући нивое његовог главног алостеричног инхибитора, малонил-ЦоА. То је зато што механизмом који је тренутно нејасан, Нрф2 негативно регулише експресију стеароил ЦоА десатуразе (СЦД) [69] и цитрат лиазе (ЦЛ) [69], [70]. Занимљиво је да нокаут или инхибиција СЦД доводи до повећане фосфорилације и активације АМП-активиране протеин киназе (АМПК) [71], [72], [73], и може се претпоставити да, у одсуству Нрф2, нивои СЦД ће се повећати, заузврат смањујући АМПК активност. Ово би могло додатно да се компликује смањеним нивоом протеина АМПК који је примећен у јетри мишева Нрф2-КО [68], што је налаз који се блиско слаже са повећаним нивоима АМПК, који су пријављени у јетри Кеап1-КД мишеви [74]. Једна последица смањене активности АМПК је ублажавање његове инхибиторне фосфорилације (на Сер79) ацетил-ЦоА карбоксилазе (АЦЦ) [75], која би могла даље да се транскрипционо регулише у одсуству Нрф2, јер је смањена активацијом Нрф2 [70 ]. Висока активност АЦЦ, у комбинацији са регулацијом ЦЛ експресије која ће повећати производњу ацетил-ЦоА, супстрата за АЦЦ, може на крају повећати ниво производа АЦЦ, малонил-ЦоА. Висок ниво малонил-ЦоА ће инхибирати ЦПТ, смањујући тако транспорт масних киселина у митохондрије. Коначно, Нрф2 позитивно регулише експресију ЦД36 [76], транслоказе која увози масне киселине кроз плазму и митохондријске мембране. Дакле, један од механизама помоћу којих Нрф2 може утицати на ефикасност митохондријске ФАО је регулисање увоза масних киселина дугог ланца у митохондрије.
Поред директне регулације транскрипције, Нрф2 такође може изменити ефикасност митохондријског ФАО својим ефектима на ћелијски редокс метаболизам. Ово може бити посебно релевантно када је активност Нрф2 ниска или је нема, услови који премештају ћелијски редокс статус у оксидовано стање. Заправо, идентификовано је неколико ФАО ензима који су осетљиви на редокс промене. Један такав ензим је врло дуголанчана ацил-ЦоА дехидрогеназа (ВЛЦАД), која више од 80% доприноси дехидрогенацији палмитоил-ЦоА у људским ткивима [77]. Занимљиво је да су Хурд и сар. [78] су показали да ВЛЦАД садржи остатке цистеина који значајно мењају своје редокс стање након излагања изолованих митохондрија срца пацова Х2О2. Поред тога, С-нитросилација мишјег јетреног ВЛЦАД на Цис238 побољшава каталитичку ефикасност ензима [79] и вероватно је да оксидација истог цистеина може имати супротан ефекат, на крају смањујући ефикасност митохондријског ФАО. Стога је могуће да, иако се нивои експресије ВЛЦАД не разликују значајно у МФ-овима ВТ, Нрф2-КО или Кеап1-КО [65], активност ензима ВЛЦАД-а може бити нижа у одсуству Нрф2 због виших нивоа РОС-а.
На основу свих ових налаза може се предложити да (слика 3): у одсуству Нрф2, нивои НАДПХ су нижи услед смањене експресије МЕ1, ИДХ1, Г6ПД и ПГД. Нивои редукованог глутатиона су такође нижи због смањене експресије ензима који учествују у његовој биосинтези и регенерацији и нижих нивоа НАДПХ потребних за конверзију оксидованог у редуковани облик глутатиона. Ниска експресија МЕ1 смањиће базен пирувата који улази у митохондрије, а гликолиза ће постати главни извор пирувата. Генерирање НАДХ је спорије, што доводи до оштећења активности комплекса И и повећане производње митохондријских РОС. Смањење ФАД-а на ФАДХ2 је такође спорије, барем делимично због мање ефикасне оксидације масних киселина, угрожавајући проток електрона из ФАДХ2 у УбК и у комплекс ИИИ. Како је УбКХ2 активатор сукцинат дехидрогеназе [80], успоравање његовог формирања може смањити ензимску активност сукцинат дехидрогеназе. Повећани нивои супероксида и водоник-пероксида могу даље инхибирати активност комплекса ИИ [81]. Нижа ефикасност оксидације масних киселина доприноси смањеној доступности супстрата за дисање митохондрија и производњу АТП у оксидативној фосфорилацији. Као компензациони механизам, гликолиза је појачана. АТП синтаза функционише обрнуто, као АТП-аза, у покушају да одржи ?? м.
Нрф2 и митохондријална биогенеза
Извештено је да, у поређењу са ВТ, јетра мишева Нрф2-КО има нижи садржај митохондрија (што је одређено односом митохондријске и нуклеарне ДНК); ово се даље смањује за 24 сата брзо и код мишева ВТ и код Нрф2-КО; за разлику од тога, иако се не разликује од ВТ у нормалним условима храњења, на пост митохондријски мишеви са високом активношћу Нрф2 не утичу [82]. Занимљиво је да суплементација Нрф2 активатором (Р) - а-липоична киселина [83], [84], [85] промовише митохондријску биогенезу у адипоцитима 3Т3-Л1 [86]. Две класе нуклеарних регулатора транскрипције играју пресудну улогу у митохондријској биогенези. Прва класа су фактори транскрипције, попут нуклеарних респираторних фактора11 и 2, који контролишу експресију гена који кодирају подјединице пет респираторних комплекса, митохондријске транслационе компоненте и хем биосинтетски ензими који су локализовани у митохондријском матриксу [88]. Пиантадоси и сар. [89] су показали да Нрф2-зависна транскрипциона регулација нуклеарног респираторног фактора 1 промовише митохондријску биогенезу и штити од цитотоксичности кардиотоксичног антрациклинског хемотерапеутског агенса доксорубицина. Супротно томе, Зханг и сар. [82] су известили да генетска активација Нрф2 не утиче на базалну експресију мРНК нуклеарног респираторног фактора 1 у мишјој јетри.
Друга класа нуклеарних регулатора транскрипције са критичним функцијама у митохондријској биогенези су коактиватори транскрипције, попут рецептора активираног пролифератором пероксисома? коактиватори (ПГЦ) 1? и 1 ?, који ступају у интеракцију са факторима транскрипције, машинама за базалну транскрипцију и РНА-спајање и ензимима који модификују хистон [88], [90], [91]. На израз породице коактиватора ПГЦ1 утичу бројни еколошки сигнали. Лечење хуманих фибробласта Нрф2 активатором сулфорафаном узрокује повећање митохондријске масе и индукцију ПГЦ1? и ПГЦ1? [92], иако у овој студији није испитивана потенцијална зависност од Нрф2. Међутим, дијабетички мишеви код којих је Нрф2 активиран хипоморфним нокдауном гена Кеап1 (дб / дб: Кеап1флок / ?: Нрф2 + / +) или поремећени (дб / дб: Кеап1флок / ?: Нрф2? /?) Имају нижи хепатички ПГЦ1? нивои експресије од контролних животиња (дб / дб: Кеап1флок / +: Нрф2 + / +) [93]. Нема разлике у нивоима мРНК за ПГЦ1? примећују се у јетри недијабетичних мишева који су или ВТ или Нрф2-КО, док су ови нивои нижи код Нрф2 прекомерно експримирајућих (Кеап1-КД и Кеап1-КО специфичних за јетру) [82]. Приметно је да 24-часовни пост повећава ниво ПГЦ1? мРНА у јетри мишева свих генотипова, али је пораст значајно већи у јетри Нрф2-КО у поређењу са мишевима који прекомерно изражавају ВТ или Нрф2. У поређењу са ВТ, мишеви Нрф2-КО који имају септичку инфекцију или акутну повреду плућа услед инфекције показују ослабљену транскрипциону регулацију нуклеарног респираторног фактора 1 и ПГЦ1? [94], [95]. Заједно, ова запажања сугеришу да је улога Нрф2 у одржавању нивоа нуклеарног респираторног фактора 1 и ПГЦ1? је сложен и постаје најистакнутији у условима стреса.
Поред експресије гена који кодирају митохондријске протеине, митохондријска биогенеза захтева и синтезу нуклеотида. Генетичка активација Нрф2 побољшава биосинтезу пурина регулацијом регулације пута пентоза-фосфата и метаболизма фолата и глутамина, посебно у брзо пролиферирајућим ћелијама (слика 2) [24]. Анализа транскриптома мутантне Дросопхиле дефицитарне за митохондријску серин / треонин протеин киназу ПТЕН-индуковану претпостављену киназу 1 (ПИНК1) показала је да митохондријска дисфункција доводи до транскрипционе регулације гена који утичу на метаболизам нуклеотида [96], сугеришући да појачана нуклеотидна биосинтеза представља механизам за заштиту од неуротоксичних последица недостатка ПИНК1. Нрф2 регулише експресију фосфорибозил пирофосфат амидотрансферазе (ППАТ), која катализује улазак у биосинтетски пут де ново пуринских нуклеотида и митохондријску метиленететрахидрофолат дехидрогеназу (МТХФД2) (слика 2). Потоњи је бифункционални ензим са активностима дехидрогеназе и циклохидролазе који је пресудан у обезбеђивању и глицина и формата као извора једнокарбонских јединица за биосинтезу пурина у брзорастућим ћелијама [2]. Стога је вероватно да би активација Нрф97 могла да буде заштитна и да преокрене митохондријску дисфункцију у недостатку ПИНК2. Заиста, фармаколошка активација Нрф1 сулфорафаном или тритерпеноидом РТА-2, обнавља ?? м и штити ћелије са недостатком ПИНК408 од токсичности допамина [1]. Иако се чини да су основни механизми сложени, ови налази указују да активност Нрф98 може утицати на биогенезу митохондрија утичући на нивое експресије критичних транскрипционих фактора и коактиватора, као и појачавањем биосинтезе нуклеотида.
Нрф2 и интегритет митохондрија
Иако директни докази нису увек доступни, постоје снажне индикације да је Нрф2 важан за интегритет митохондрија, посебно у условима оксидативног стреса. Митохондрији изоловани из мозга и јетре пацова којима је дата појединачна доза Нрф2 активатора сулфорафана отпорни су на отварање прелазне поре митохондријалне пермеабилности (мПТП) изазване оксидантом терц-бутилхидропероксидом [99], [100]. МПТП, комплекс који омогућава унутрашњој мембрани митохондрија да постане пропусна за молекуле масе до 1500 Да, недавно је утврђено да се формира из димера Ф0Ф1-АТП синтазе [101]. Отпор посредством сулфорафана на отварање мПТП корелира са повећаном антиоксидативном одбраном, а нивои митохондријског ГСХ, глутатион пероксидазе 1, јабучни ензим 3 и тиоредоксин 2 повећани су у фракцијама митохондрија изолованим од животиња третираних сулфорафаном [100].
Оштећење митохондријских протеина и оштећење дисања изазвано електрофилним продуктом пероксидације липида 4-хидрокси-2-ноненал ослабљени су у митохондријима изолованим из мождане коре мишева третираних сулфорафаном [102]. У бубрежним епителним ћелијама пацова и у бубрезима, сулфорафан штити од токсичности изазване цисплатином и гентамицином и губитка ?? м [103], [104]. Заштита од панела оксиданата (супероксид, водоник-пероксид, пероксинитрит) и електрофила (4-хидрокси-2-ноненал и акролеин) и повећање митохондријске антиоксидативне одбране примећене су и током лечења глатких мишићних ћелија аорте пацова сулфорафаном [105 ]. У моделу акутне повреде бубрега изазване контрастом, недавно је показано да исхемијско предусловљавање екстремитета има заштитне ефекте, укључујући инхибицију отварања мПТП-а и митохондријског отока, активацијом Нрф2 услед инхибиције ГСК3? [106].
Митофагија, процес којим дисфункционалне митохондрије селективно прогутају аутофагосоми и достављају их у лизосоме да би их ћелија разградила и рециклирала, од суштинског је значаја за митохондријску хомеостазу [107], [108]. Иако није утврђена узрочна веза између Нрф2 и митофагије, постоје докази да би фактор транскрипције могао бити важан у контроли квалитета митохондрија играјући улогу у митофагији. Ово би могло бити посебно видљиво у условима оксидативног стреса. Дакле, у моделу сепсе, пораст нивоа аутофагосомског маркера МАП1 лаки ланац 3-ИИ (ЛЦ3-ИИ) и теретног протеина п62 за 24 сата након инфекције сузбија се у Нрф2-КО у поређењу са ВТ мишевима [109] . Недавно је откривен индуктор митофагије са малим молекулом (звани индуктор митофагије посредством п62, ПМИ); ово једињење 1,4-дифенил-1,2,3-триазола је првобитно дизајнирано као Нрф2 активатор који нарушава интеракцију транскрипционог фактора са Кеап1 [110]. Слично ћелијама у којима је Нрф2 генетски регулисан (Кеап1-КД или Кеап1-КО), ћелије изложене ПМИ имају веће мировање ?? м. Важно је да се повећање локализације митохондрија ЛЦ3, које се примећује након ПМИ третмана ВТ ћелија, не дешава у Нрф2-КО ћелијама, што указује на умешаност Нрф2.
На крају, ултраструктурна анализа делова јетре открила је присуство отечених митохондрија са смањеним кристама и поремећеним мембранама у хепатоцитима мишева Нрф2-КО, али не и ВТ, који су храњени храном са пуно масти током 24 недеље; посебно, ове јетре показују јасне доказе оксидативног стреса и упале [68]. Може се закључити да Нрф2 има пресудну улогу у одржавању интегритета митохондрија у условима оксидативног и инфламаторног стреса.
Сулфорафан и његови ефекти на рак, морталитет, старење, мозак и понашање, болести срца и друго
Изотиоцијанати су нека од најважнијих биљних једињења која можете добити у исхрани. У овом видеу радим за њих најсвеобухватнији случај који је икада направљен. Кратак распон пажње? Пређите на своју омиљену тему кликом на једну од временских тачака у наставку. Комплетна временска линија испод.
Кључни делови:
00:01:14 - Рак и смртност
00:19:04 - Старење
00:26:30 - Мозак и понашање
00:38:06 - Коначни приказ
00:40:27 - Доза
Комплетна временска линија:
00:00:34 - Увођење сулфорафана, главни фокус видео снимка.
00:01:14 - Потрошња крстастих поврћа и смањење смртности од свих узрока.
00:02:12 - Ризик од рака простате.
00:02:23 - Ризик од рака бешике.
00:02:34 - Рак плућа код пушача ризикује.
00:02:48 - Ризик од рака дојке.
00:03:13 - Хипотетично: шта ако већ имате рак? (интервенцијски)
00:03:35 – Уверљиви механизам који покреће асоцијативне податке о раку и морталитету.
00:04:38 - Сулфорафан и рак.
00:05:32 – Докази на животињама показују снажан ефекат екстракта клице броколија на развој тумора бешике код пацова.
00:06:06 - Ефекат директне суплементације сулфорафана код пацијената са раком простате.
00:07:09 - Биоакумулација метаболита изотиоцијаната у стварном ткиву дојке.
00:08:32 - Инхибиција матичних ћелија рака дојке.
00:08:53 - Лекција из историје: брассицас је утврдио да има здравствена својства чак и у старом Риму.
00:09:16 - Сулфорафанова способност да појача излучивање канцерогена (бензен, акролеин).
00:09:51 - НРФ2 као генетски прекидач преко елемената антиоксидативног одговора.
00:10:10 - Како активација НРФ2 појачава излучивање канцерогена преко глутатиона-С-коњугата.
00:10:34 - Кељ у Бриселу повећава глутатион-С-трансферазу и смањује оштећење ДНК.
00:11:20 - Напитак од клице брокуле повећава излучивање бензела за 61%.
00:15:45 - Конзумација крстастих поврћа и смртност од срчаних болести.
00:16:55 - Прашак клице брокуле побољшава липиде у крви и укупан ризик од болести срца код дијабетичара типа 2.
00:19:04 – Почетак секције старења.
00:19:21 – Исхрана обогаћена сулфорафаном продужава животни век буба од 15 до 30% (у одређеним условима).
00:20:34 - Значај слабе упале за дуговечност.
00:22:05 - Чини се да крстасто поврће и прах клица брокуле смањују широк спектар маркера упале код људи.
00:23:40 - Осврт на видео запис: рак, делови за старење
00:24:14 - Студије на мишевима сугеришу да сулфорафан може побољшати адаптивну имунолошку функцију у старости.
00:25:18 – Сулфорафан је побољшао раст косе на мишјем моделу ћелавости. Слика на 00:26:10.
00:26:30 - Почетак одељења за мозак и понашање.
00:27:18 - Ефекат екстракта клице брокуле на аутизам.
00:27:48 - Ефекат глукорафанина на шизофренију.
00:28:17 - Почетак дискусије о депресији (вероватни механизам и студије).
00:31:21 – Студија на мишу користећи 10 различитих модела депресије изазване стресом показује да је сулфорафан сличан ефикасан као флуоксетин (прозак).
00:32:00 - Студија показује да је директно уношење глукорафанина код мишева слично ефикасно у спречавању депресије из модела стреса социјалног пораза.
00:34:43 - Почетак одељења са трауматском повредом мозга.
00:35:01 - Сулфорафан убризган непосредно након ТБИ побољшава меморију (студија миша).
00:35:55 - Сулфорафан и неуронска пластичност.
00:36:32 – Сулфорафан побољшава учење у моделу дијабетеса типа ИИ код мишева.
00:37:19 – Сулфорафан и Дуцхенне мишићна дистрофија.
00:37:44 - Инхибиција миостатина у мишићним сателитским ћелијама (ин витро).
00:38:06 - Касни видео-приказ: смртност и рак, оштећење ДНК, оксидативни стрес и упале, излучивање бензена, кардиоваскуларне болести, дијабетес типа ИИ, ефекти на мозак (депресија, аутизам, шизофренија, неуродегенерација), пут НРФ2.
00:40:27 - Размишљања о проналажењу дозе клица брокуле или сулфорафана.
00:41:01 - Анегдоте о ницању код куће.
00:43:14 - О температурама кувања и активности сулфорафана.
00:43:45 - Конверзија бактерија у цревима сулфорафана из глукорафанина.
00:44:24 - Додаци делују боље у комбинацији са активном мирозиназом из поврћа.
00:44:56 - Технике кувања и крстасто поврће.
00:46:06 - Изотиоцијанати као гоитрогени.
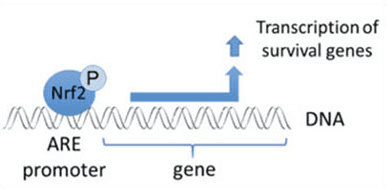
Нрф2 је фактор транскрипције који игра важну улогу у ћелијском антиоксидативном одбрамбеном систему људског тела. Елемент који реагује на антиоксидант, или АРЕ, је регулаторни механизам гена. Многа истраживачка истраживања су показала да Нрф2, или фактор 2 повезан са НФ-Е2, регулише широк спектар АРЕ покретаних гена у неколико врста ћелија. Такође је утврђено да Нрф2 игра суштинску улогу у ћелијској заштити и антиканцерогености, што показује да Нрф2 може бити ефикасан третман у лечењу неуродегенеративних болести и карцинома за које се верује да су узроковани оксидативним стресом. Др Алек Јименез ДЦ, ЦЦСТ Инсигхт
Завршне напомене
Иако многа питања и даље остају отворена, доступни експериментални докази јасно указују да је Нрф2 важан играч у одржавању митохондријске хомеостазе и структурног интегритета. Ова улога постаје посебно критична у условима оксидативног, електрофилног и инфламаторног стреса када способност појачаног регулисања Нрф2 посредованих цитопротективних одговора утиче на целокупно здравље и опстанак ћелије и организма. Улога Нрф2 у функцији митохондрија представља још један слој широких цитопротективних механизама које оркестрира овај фактор транскрипције. Како многа људска патолошка стања имају оксидативни стрес, упале и митохондријску дисфункцију као кључне компоненте њихове патогенезе, фармаколошка активација Нрф2 обећава за превенцију и лечење болести. Свеобухватно разумевање прецизних механизама којима Нрф2 утиче на функцију митохондрија неопходан је за рационалан дизајн будућих клиничких испитивања и може понудити нове биомаркере за праћење терапијске ефикасности.
Сврха горњег чланка била је да се дискутује, као и да се демонстрира, нова улога Нрф2 у функцији митохондрија. Нрф2, или нуклеарни фактор фактор који се односи на еритроид 2, је нови регулатор ћелијске отпорности на оксиданте који може допринети оксидативном стресу, утичући на ћелијску функцију и довести до развоја токсичности, хроничних болести, па чак и рака. Док производња оксиданата у људском телу може служити у разне сврхе, укључујући деобу ћелија, упалу, имунолошку функцију, аутофагију и одговор на стрес, неопходно је контролисати њихову прекомерну производњу како би се спречили здравствени проблеми. Обим наших информација је ограничен на киропрактику и здравствене проблеме кичме. Да бисте разговарали о овој теми, слободно питајте др Хименеза или нас контактирајте на�915-850-0900 .
Бол у леђима�један од најчешћих узрока инвалидитета и пропуштених дана на послу широм света. Бол у леђима приписује другом најчешћем разлогу посета лекару, надмашују га само инфекције горњих дисајних путева. Отприлике 80 процената популације искусиће бол у леђима бар једном током свог живота. Кичма је сложена структура коју чине кости, зглобови, лигаменти и мишићи, између осталих меких ткива. Због тога, повреде и/или отежана стања, као што је�хернија дискова, може на крају довести до симптома болова у леђима. Спортске повреде или повреде у саобраћајним незгодама често су најчешћи узрок болова у леђима, међутим, понекад најједноставнији покрети могу имати болне последице. На срећу, алтернативне опције лечења, као што је киропрактичка нега, могу помоћи у ублажавању болова у леђима коришћењем подешавања кичме и ручних манипулација, што на крају побољшава ублажавање болова. �
Нрф2 подржава активацију групе антиоксидативних и детоксикационих ензима и гена који штите људско тело од ефеката здравствених проблема повезаних са повећаним нивоима оксидативног стреса, као што је Алцхајмерова болест. Доказано је да разне природне супстанце активирају пут Нрф2, који може помоћи у управљању симптомима неуродегенеративних болести. Сврха чланка у наставку је да се расправља о кључној улози Нрф2 узрокованој хроничном упалом.
Апстрактан
Упала је најчешћа карактеристика многих хроничних болести и компликација, док игра кључну улогу у канцерогенези. Неколико студија је показало да Нрф2 доприноси антиинфламаторном процесу оркестрирајући регрутовање инфламаторних ћелија и регулишући експресију гена кроз елемент антиоксидативног одговора (АРЕ). Кеап1 (Келцх-лике ЕЦХ-ассоциатед протеин)/Нрф2 (НФ-Е2 п45-везан фактор 2)/АРЕ сигнални пут углавном регулише експресију антиинфламаторних гена и инхибира напредовање упале. Стога је идентификација нових Нрф2 зависних антиинфламаторних фитокемикалија постала кључна тачка у откривању лекова. У овом прегледу разматрамо чланове Кеап1/Нрф2/АРЕ сигналног пута и његове низводне гене, ефекте овог пута на животињске моделе инфламаторних болести и преслушавање са НФ-?Б путем. Поред тога, такође разговарамо о регулацији инфламасома НЛРП3 од стране Нрф2. Осим тога, резимирамо тренутни сценарио развоја антиинфламаторних фитокемикалија и других који посредују у Нрф2/АРЕ сигналном путу.
Упала је сложен процес који се јавља када су ткива инфицирана или повређена штетним стимулансима као што су патогени, оштећења или иританси. Имунске ћелије, крвни судови и молекуларни медијатори су укључени у овај заштитни одговор [1]. Упала је такође патолошки феномен повезан са разним болесним стањима изазваним углавном физичким, хемијским, биолошким и психолошким факторима. Циљ инфламације је да ограничи и елиминише узроке оштећења ћелија, очисти и/или апсорбује некротичне ћелије и ткива и започне поправку ткива. Разликују се два различита облика упале: акутни и хронични. Акутна упала је самоограничавајућа и корисна за домаћина, али продужена хронична упала је уобичајена карактеристика многих хроничних болести и компликација. Директна инфилтрација многих мононуклеарних имуних ћелија као што су моноцити, макрофаги, лимфоцити и плазма ћелије, као и производња инфламаторних цитокина, доводе до хроничне упале. Познато је да хронична упала игра кључну улогу у канцерогенези [2]. Генерално, и про- и антиинфламаторни сигнални путеви међусобно делују у нормалном инфламаторном процесу.
У патолошком инфламаторном процесу, прво се активирају мастоцити, моноцити, макрофаги, лимфоцити и друге имуне ћелије. Затим се ћелије регрутују на место повреде, што доводи до стварања реактивних врста кисеоника (РОС) које оштећују макромолекуле укључујући ДНК. У исто време, ове инфламаторне ћелије такође производе велике количине медијатора упале као што су цитокини, хемокини и простагландини. Ови медијатори даље регрутују макрофаге на локализована места упале и директно активирају вишеструке каскаде трансдукције сигнала и факторе транскрипције повезане са упалом. НФ-?Б (нуклеарни фактор капа Б), МАПК (митоген-активирана протеин киназа) и ЈАК (јанус киназа)-СТАТ (претварачи сигнала и активатори транскрипције) сигнални путеви су укључени у развој класичног пута упале [3], [4], [5]. Претходне студије су откриле да фактор транскрипције Нрф2 (НФ-Е2 п45-везан фактор 2) регулише експресију ензима за детоксикацију фазе ИИ укључујући НАДПХ, НАД(П)Х кинон оксидоредуктазу 1, глутатион пероксидазу, феритин, хем оксигеназу-1 (ХО -1), и антиоксидативних гена који својим антиинфламаторним дејством штите ћелије од разних повреда, утичући на ток болести [6], [7], [8].
Узимајући у обзир ове изузетне налазе, развој циљаних терапијских лекова за инфламаторне болести путем сигналних путева привукао је велико интересовање последњих година. У овом прегледу сумирамо истраживање сигналног пута Кеап1 (ЕЦХ-повезан протеин сличан Келцху)/Нрф2 (фактор 2 повезан са НФ-Е45 п2)/АРЕ (антиоксидативни елемент одговора) сигналног пута.
Структура и регулација Нрф2
Кеап1-зависна Нрф2 регулатива
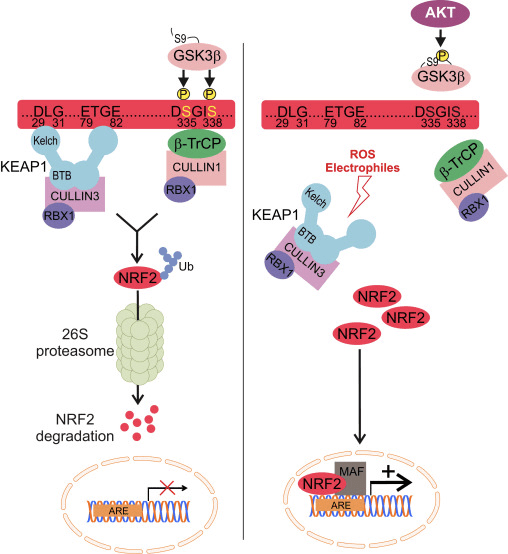
Нрф2 припада подфамилији Цап �н� Цоллар (ЦНЦ) и састоји се од седам функционалних домена, Нех (Нрф2-ЕЦХ хомологија) 1 до Нех7 [9], [10]. Нех1 је ЦНЦ-бЗИП домен који омогућава Нрф2 да се хетеродимеризује са малим мускулоапонеуротичним фибросаркомом (Маф) протеином, ДНК и другим транскрипционим партнерима, као и да формира нуклеарни комплекс са ензимом који коњугира убиквитин УбцМ2 [11], [12]. Нех2 садржи два важна мотива позната као ДЛГ и ЕТГЕ, који су неопходни за интеракцију између Нрф2 и његовог негативног регулатора Кеап1 [13], [14].
Кеап1 је адаптер супстрата за Е3 убиквитин лигазу засновану на кулину, који инхибира транскрипциону активност Нрф2 путем убиквитинације и протеазомалне деградације у нормалним условима [15], [16], [17]. КЕЛЦХ домени Кеап1 хомодимера се везују за ДЛГ и ЕТГЕ мотиве Нрф2-Нех2 домена у цитосолу, где ЕТГЕ делује као шарка са већим афинитетом, а ДЛГ делује као квачица [18]. Под оксидативним стресом или након излагања активаторима Нрф2, Нрф2 се одваја од Кеап1 везивања због тиолне модификације Кеап1 цистеинских остатака што на крају спречава убиквитинацију Нрф2 и протеазомску деградацију [19]. Затим се Нрф2 транслоцира у језгро, хетеродимеризује са малим Маф протеинима и трансактивира АРЕ батерију гена (слика 1А). Карбокси-терминал Нех3 делује као трансактивациони домен интеракцијом са коактиватором транскрипције познатим као ЦХД6 (протеин који везује ДНК хромо-АТПазе/хеликазе) [20]. Нех4 и Нех5 такође делују као домени трансактивације, али се везују за други транскрипциони коактиватор познат као ЦБП (цАМП-респонсе-елемент-биндинг протеин-биндинг протеин) [21]. Штавише, Нех4 и Нех5 ступају у интеракцију са нуклеарним кофактором РАЦ3/АИБ1/СРЦ-3, што доводи до побољшане експресије АРЕ гена циљаног на Нрф2 [22]. Нех5 има нуклеарни извозни сигнал осетљив на редокс који је кључан за регулацију и ћелијску локализацију Нрф2 [23].
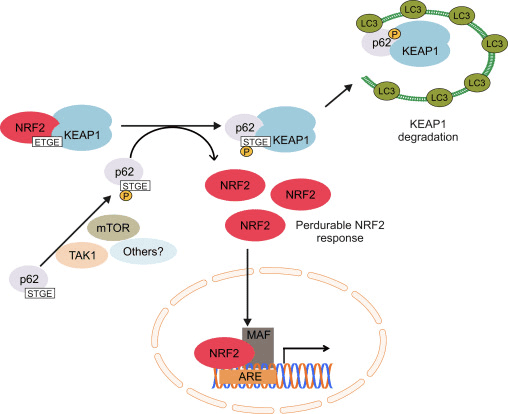
Слика 1 Кеап1 зависна и независна регулација Нрф2. (А) У базалним условима, Нрф2 је одвојен са Кеап1 са своја два мотива (ЕТГЕ и ДЛГ) што доводи до убиквитинације посредоване ЦУЛ3 праћеном деградацијом протеасома. Под оксидативним стресом, Нрф2 се дисоцира од Кеап1, транслоцира у језгро и активира АРЕ-генску батерију. (Б) ГСК3 фосфорилише Нрф2 и то олакшава препознавање Нрф2 помоћу ?-ТрЦП за убиквитинацију посредовану ЦУЛ1 и каснију деградацију протеазома. (Ц) п62 је одвојен са Кеап1, што доводи до његове аутофагне деградације, ослобађања Нрф2 и повећане сигнализације Нрф2.
Кеап1-Индепендент Нрф2 Регулатион
Нови докази су открили нови механизам регулације Нрф2 који је независан од Кеап1. Серином богат Нех6 домен Нрф2 игра кључну улогу у овој регулацији везивањем са своја два мотива (ДСГИС и ДСАПГС) за протеин који садржи понављање ?-трансдуцина (?-ТрЦП) [24]. ?-ТрЦП је супстратни рецептор за комплекс убиквитин лигазе Скп1�Цул1�Рбк1/Роц1 који циља на Нрф2 за убиквитинацију и протеазомалну деградацију. Гликоген синтаза киназа-3 је кључни протеин укључен у Кеап1 независну Нрф2 стабилизацију и регулацију; он фосфорилише Нрф2 у Нех6 домену да би олакшао препознавање Нрф2 помоћу ?-ТрЦП и накнадну деградацију протеина [25] (слика 1Б).
Други Нрф2 регулатори
Друга линија доказа је открила неканонски пут п62-зависне активације Нрф2 у којој п62 секвестрира Кеап1 до аутофагне деградације која на крају доводи до стабилизације Нрф2 и трансактивације гена зависних од Нрф2 [26], [27], [ 28], [29] (слика 1Ц).
Акумулирани докази сугеришу да неколико миРНА игра важну улогу у регулацији активности Нрф2 [30]. Сангокоиа ет ал. [31] је показао да миР-144 директно смањује активност Нрф2 у ћелијској линији лимфобласта К562, примарним хуманим еритроидним прогенитор ћелијама и ретикулоцитима српастих ћелија. Још једна занимљива студија у ћелијама епитела дојке код људи показала је да миР-28 инхибира Нрф2 путем Кеап1 независног механизма [32]. Слично, миРНА као што су миР-153, миР-27а, миР-142-5п и миР144 смањују експресију Нрф2 у ћелијској линији неурона СХ-СИ5И [33]. Сингх ет ал. [34] су показали да ектопична експресија миР-93 смањује експресију Нрф2 регулисаних гена у 17а-естрадиолом (Е2) индукованом моделу карциногенезе дојке код пацова.
Недавно откриће из наше лабораторије идентификовало је ендогени инхибитор Нрф2 познат као ретиноични Кс рецептор алфа (РКСР?). РКСР? је нуклеарни рецептор, интерагује са Нех7 доменом Нрф2 (остаци аминокиселина 209×316) преко свог ДНК-везујућег домена (ДБД), и специфично инхибира активност Нрф2 у језгру. Штавише, пријављени су и други нуклеарни рецептори као што су рецептор-а активиран пролифератором пероксизома, ЕРа, рецептор-а повезан са естрогеном и глукокортикоидни рецептори као ендогени инхибитори активности Нрф2 [9], [10].
Анти-инфламаторна улога Нрф2/ХО-1 осе
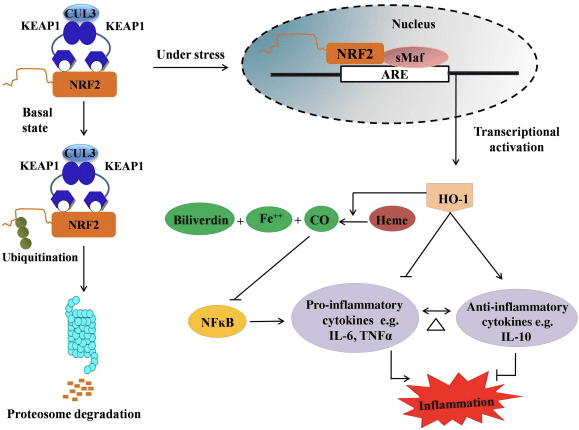
ХО-1 је индуцибилна изоформа и ензим који ограничава брзину који катализује разградњу хема у угљен моноксид (ЦО) и слободно гвожђе, и биливердина у билирубин. Ензимска деградација проинфламаторног слободног хема као и производња антиинфламаторних једињења као што су ЦО и билирубин играју главну улогу у одржавању заштитних ефеката ХО-1 (слика 2).
Слика 2 Преглед Нрф2/ХО-1 пута. У базалним условима, Нрф2 се везује за свој репресор Кеап1 што доводи до убиквитинације праћене деградацијом протеасома. Током оксидативног стреса, слободни Нрф2 се транслоцира у језгро, где се димеризује са члановима мале породице Маф и везује се за АРЕ гене као што је ХО-1. Повећани ХО-1 катализује хем у ЦО, билирубин и слободно гвожђе. ЦО делује као инхибитор НФ-?Б пута што доводи до смањене експресије проинфламаторних цитокина, док билирубин делује и као антиоксиданс. Даље, ХО-1 директно инхибира проинфламаторне цитокине као и активирање антиинфламаторних цитокина, што доводи до балансирања инфламаторног процеса.
Нрф2 индукује ХО-1 ген повећањем експресије мРНА и протеина и један је од класичних Нрф2 регулисаних гена који се широко користи у бројним ин витро и ин виво студијама. Неколико студија је показало да ХО-1 и његови метаболити имају значајне антиинфламаторне ефекте посредоване Нрф2. Повећање експресије ХО-1 које је посредовано активираним Нрф2 доводи до инхибиције НФ?Б сигнализације резултира смањеном повредом цревне слузокоже и дисфункцијом чврстог споја код модела трансплантације јетре мушког Спрагуе-Давлеи пацова [35]. Повећање експресије ХО-2 зависне од Нрф1 може заштитити миобласте Ц2Ц12 добијене од миша од цитотоксичности Х2О2 [36]. ХО-2 зависан од Нрф1 има утицај на инфламаторне одговоре посредоване липополисахаридом (ЛПС) у РАВ264.7- или макрофагима пенастих ћелија добијених из перитонеалног макрофага миша. Активност Нрф2 десензибилизира фенотип макрофага пенастих ћелија и спречава неумерено запаљење макрофага, који играју важну улогу у прогресији атеросклерозе [37]. Оса Нрф2/ХО-1 утиче на микроглијалне ћелије БВ2 миша индуковане ЛПС и ХТ22 ћелије хипокампуса миша, са утицајем на неуроинфламацију. Повећање експресије ХО-1 путем Нрф2 пута у микроглијалним ћелијама БВ2 миша које бране ћелијску смрт ХТ22 ћелија хипокампуса миша [38]. Штавише, хибридни молекули на бази кобалта (ХИЦО) који комбинују индуктор Нрф2 са ослобађачем угљен моноксида (ЦО) повећавају експресију Нрф2/ХО-1, ослобађају ЦО и испољавају антиинфламаторну активност ин витро. ХИЦО такође повећавају регулацију ткива ХО-1 и испоручују ЦО у крв након примене ин виво, подржавајући њихову потенцијалну употребу против упалних стања [39]. Повећана регулација Нрф2/ХО-1 смањује упалу повећањем ефероцитне активности мишјих макрофага третираних таурин хлораминима [40]. Све у свему, горе објашњени експериментални модели су открили да Нрф2/ХО-1 оса игра главну улогу у антиинфламаторној функцији, што сугерише да је Нрф2 терапеутска мета код болести повезаних са упалом.
Поред тога, нуспродукти ХО-1 као што је ЦО, билирубин, делује као моћан антиоксиданс током оксидативног стреса и оштећења ћелија [41], [42]; сузбија аутоимуни енцефаломијелитис и хепатитис [43], [44]; и штити мишеве и пацове од ендотоксичног шока спречавањем стварања иНОС и НО [45], [46], [47]. Штавише, билирубин смањује ендотелну активацију и дисфункцију [48]. Занимљиво је да билирубин смањује трансмиграцију ендотелних леукоцита преко адхезионог молекула-1 [49]. Ове специфичне референце указују на то да не само да ХО-1 делује као моћно антиинфламаторно средство, већ и његови метаболити.
Инфламаторни медијатори и ензими инхибирани Нрф2
Цитокини и хемокини
Цитокини су протеини ниске молекуларне тежине и полипептиди које луче различите ћелије; регулишу раст ћелија, диференцијацију и имунолошку функцију, а укључени су у запаљење и зарастање рана. Цитокини укључују интерлеукине (ИЛ), интерфероне, фактор некрозе тумора (ТНФ), фактор стимулације колонија, хемокине и факторе раста. Неки цитокини се рачунају као проинфламаторни медијатори, док други имају антиинфламаторне функције. Изложеност оксидативном стресу доводи до прекомерне производње цитокина што изазива оксидативни стрес у циљним ћелијама. Неколико проинфламаторних цитокина се прекомерно производи када се НФ-?Б активира оксидативним стресом. Штавише, проинфламаторни оксидативни стрес изазива даљу активацију НФ-?Б и прекомерну производњу цитокина. Активација Нрф2/АРЕ система игра важну улогу у нарушавању овог циклуса. Хемокини су породица малих цитокина, чија је главна улога да усмеравају миграцију инфламаторних ћелија. Они функционишу углавном као хемоатрактанти за леукоците, моноците, неутрофиле и друге ефекторске ћелије.
Пријављено је да активација Нрф2 спречава ЛПС-индуковану транскрипциону регулацију проинфламаторних цитокина, укључујући ИЛ-6 и ИЛ-1? [50]. ИЛ-1? и производња ИЛ-6 је такође повећана у Нрф2?/? мишеви са колитисом изазваним декстран сулфатом [51], [52]. Нрф2 инхибира производњу низводног ИЛ-17 и других инфламаторних фактора Тх1 и Тх17 и сузбија процес болести у експерименталном моделу мултипле склерозе, аутоимуног енцефалитиса [53]. Нрф2 зависни антиоксидативни гени ХО-1, НКО-1, Гцлц и Гцлм блокирају ТНФ-а, ИЛ-6, протеин хемоатрактанта моноцита-1 (МЦП1), инфламаторни протеин-2 макрофага (МИП2) и инфламаторни протеин посредници. Али у случају Нрф2-нокаут мишева, антиинфламаторни ефекат се не јавља [54]. Перитонеални неутрофили Нрф2-нокаут мишева третираних ЛПС-ом имају значајно више нивое цитокина (ТНФ-а и ИЛ-6) и хемокина (МЦП1 и МИП2) него ћелије дивљег типа (ВТ) [54]. Ин витро, пренос гена Нрф2 на ћелије глатких мишића аорте људи и зеца потискује секрецију МЦП1 [8], [55], а експресија ХО-2 зависна од Нрф1 потискује НФ-?-стимулисане НФ-?Б и МЦП-1 секреција у ендотелним ћелијама људске пупчане вене [56]. Ови налази наговештавају да, као одговор на инфламаторне стимулусе, појачана регулација Нрф2 сигнализације инхибира прекомерну производњу проинфламаторних цитокина и хемокина, као и ограничавање активације НФ-?Б.
Молекули ћелијске адхезије
Молекули ћелијске адхезије (ЦАМ) су протеини који се везују за ћелије или са екстрацелуларним матриксом. Смештени на површини ћелије, они су укључени у препознавање ћелија, активацију ћелије, трансдукцију сигнала, пролиферацију и диференцијацију. Међу ЦАМ-овима, ИЦАМ-1 и ВЦАМ-1 су важни чланови суперфамилије имуноглобулина. ИЦАМ-1 је присутан у ниским концентрацијама у мембранама леукоцита и ендотелних ћелија. Након стимулације цитокина, концентрација се значајно повећава. ИЦАМ-1 може бити индукован ИЛ-1 и ТНФ и експримиран је васкуларним ендотелом, макрофагима и лимфоцитима. То је лиганд за интегрин, рецептор који се налази на леукоцитима. Када се активира ИЦАМ-1-интегрински мост, леукоцити се везују за ендотелне ћелије и затим мигрирају у субендотелна ткива [57]. ВЦАМ-1 посредује у адхезији лимфоцита, моноцита, еозинофила и базофила на васкуларни ендотел и доприноси регрутовању леукоцита, што на крају доводи до оштећења ткива услед оксидативног стреса. Нрф2 инхибира промоторску активност ВЦАМ-1 [58]. Низводни ген ХО-2 регулисан Нрф1 може утицати на експресију Е-селектина и ВЦАМ-1, адхезионих молекула повезаних са ендотелним ћелијама [59]. Плућна експресија неколико ЦАМ-ова као што су ЦД-14, ТРЕМ1, СЕЛЕ, СЕЛП и ВЦАМ-1 је значајно већа у Нрф2?/? мишева него код Нрф2+/+ мишева [60]. Нрф2 у људским ендотелним ћелијама аорте сузбија експресију ВЦАМ-1 изазвану ТНФ-а и омета ТНФ-а-индуковану моноцитну У937 ћелијску адхезију [8]. Прекомерна експресија Нрф2 такође инхибира експресију гена ВЦАМ-1 изазвану ТНФ-а у хуманим микроваскуларним ендотелним ћелијама [61]. Утврђено је да природни антиоксидант 3-хидроксиантранилна киселина (ХА), један од метаболита л-триптофана који се формира ин виво дуж метаболичког пута познатог као кинуренин током упале или инфекције, индукује експресију ХО-1 и стимулише Нрф2 у пупчаном каналу људи. ендотелне ћелије вене (ХУВЕЦс). Експресија ХО-2 зависна од Нрф1 индукована ХА инхибира секрецију МЦП-1, експресију ВЦАМ-1 и активацију НФ-кБ повезану са васкуларним повредама и упалом код атеросклерозе [56]. Анти-пролиферативни и антиинфламаторни синтетички дериват халкона 2?,4?,6?-трис (метоксиметокси) халкон инхибира ИЦАМ-1, проинфламаторни цитокин ИЛ-1? и ТНФ-? експресија у ткиву дебелог црева код мишева третираних тринитробензен сулфонском киселином [62]. Повећање регулације Нрф2 инхибира ТНФ-а-индуковану експресију ИЦАМ-1 у епителним ћелијама пигмента ретине људи третираних ликопеном [63]. Све ове студије сугеришу да Нрф2 игра кључну улогу у инфламаторном процесу тако што регулише миграцију и инфилтрацију инфламаторних ћелија у упаљено ткиво.
Матрикс металопротеиназе (ММП)
ММП су широко присутни у екстрацелуларном матриксу и укључени су у физиолошке и патолошке процесе као што су пролиферација ћелија, миграција, диференцијација, зарастање рана, ангиогенеза, апоптоза и метастазе тумора. Пријављено је да осовина Нрф2/ХО-1 инхибира ММП-9 у макрофагима и ММП-7 у хуманим интестиналним епителним ћелијама, и то је корисно у лечењу инфламаторне болести црева [62], [64]. Оштећење коже изазвано УВ зрачењем је теже код Нрф2-нокаута него код ВТ мишева и ниво ММП-9 је значајно виши, што указује да Нрф2 смањује експресију ММП-9. Стога се сматра да је Нрф2 заштитник од УВ зрачења [65]. Друга студија је такође известила да је смањена транскрипциона активација ММП-9 у инвазији туморских ћелија и инфламацији регулисана инхибицијом НФ-кБ сигналног пута [66]. Код трауматског оштећења кичмене мождине, НФ-кБ сигнални пут такође учествује у регулисању нивоа мРНК ММП-9 [67]. Стога, код инфламације на регулацију ММП директно утиче Нрф2 пут или индиректно преко НФ-?Б пута под утицајем Нрф2.
Циклооксигеназа-2 (ЦОКС2) и индуцибилна синтаза азот-оксида (ИНОС)
Серија експеримената на Нрф2-нокаут мишевима је показала његову кључну улогу у упали и регулацији проинфламаторних гена као што су ЦОКС-2 и иНОС. По први пут, Кхор ет ал. пријавили повећану експресију проинфламаторних цитокина као што су ЦОКС-2 и иНОС у ткивима дебелог црева Нрф2?/? мишеви у поређењу са ВТ Нрф2+/+ мишевима, што указује да Нрф2 потискује њихову активност [51]. Други извештај о претходном третману сулфорафаном, једним од добро познатих активатора Нрф2 присутних у поврћу крсташа, показао је његов антиинфламаторни ефекат инхибиције експресије ТНФ-а, ИЛ-1?, ЦОКС-2 и иНОС на обе мРНА и нивои протеина у примарним перитонеалним макрофагима из Нрф2+/+ мишева у поређењу са онима из Нрф2?/? мишеви [68]. Слично, хипокампус Нрф2-нокаут мишева са упалом изазваном ЛПС такође показује већу експресију маркера упале као што су иНОС, ИЛ-6 и ТНФ-? него ВТ мишеви [69]. Слично, Нрф2-нокаут мишеви су преосетљиви на оксидативни стрес изазван 1-метил-4-фенил-1,2,3,6-тетрахидропиридин, као и показују повећане нивое мРНА и протеина маркера запаљења као што су ЦОКС-2, иНОС , ИЛ-6 и ТНФ-? [70]. Штавише, јетре из Нрф2?/? мишеви који су подвргнути исхрани са недостатком метионина и холина имају ~ 5 пута већу експресију мРНК Цок2 и иНОС од оних код ВТ мишева на истој исхрани, што указује на антиинфламаторну улогу Нрф2 [71]. Недавно су Ким ет ал. показали су да фитокемијска етил пируват испољава своје антиинфламаторне и антиоксидативне ефекте смањујући експресију иНОС-а путем Нрф2 сигнализације у БВ2 ћелијама. Они су показали да етил пируват индукује нуклеарну транслокацију Нрф2, која на крају инхибира интеракцију између п65 и п300, што доводи до смањене експресије иНОС [72]. Штавише, аналог карбазола ЛЦИ-2-ЦХО активира Нрф2 и изазива његову нуклеарну транслокацију, што доводи до супресије експресије ЦОКС2 и иНОС [73] у ћелијама глатких мишића крвних судова аорте пацова.
Парадоксална улога Нрф2 у регулацији активности ифламасома НЛРП3
Породица НЛР, пирински домен који садржи 3 (НЛРП3) инфламазом је мултипротеински комплекс који функционише као рецептор за препознавање патогена (ПРР) и препознаје широк спектар микробних сигнала оксидативног стреса као што су молекуларни обрасци повезани са патогеном (ПАМП), оштећење- повезане молекуле молекуларног узорка (ДАМП) и РОС [74]. Активирани инфламазом НЛРП3 посредује у цепању каспазе-1 и секрецији проинфламаторног цитокина интерлеукина-1? (ИЛ-1?) који на крају индукује процес ћелијске смрти познат као пироптоза који штити домаћине од широког спектра патогена [75]. Међутим, аберантна активација инфламазома је повезана са болестима погрешног савијања протеина као што су преносиве спонгиформне енцефалопатије, Алцхајмерова болест, Паркинсонова болест, као и дијабетес типа 2 [76], рак [77], гихт и атеросклероза [78].
Недавно запажање групе Ронг Ху о повезаности Нрф2 са негативном регулацијом инфламазома открило је да Нрф2 индукује експресију НКО1 која доводи до инхибиције активације НЛРП3 инфламасома, цепања каспазе-1 и ИЛ-1? генерације у макрофагима. Штавише, добро познати активатор Нрф2, терц-бутилхидрокинон (тБХК) негативно је регулисао транскрипцију НЛРП3 активирањем АРЕ на Нрф2 зависан начин [79]. Поред горе наведеног запажања, иста група је такође откривена да диметил фумарат (ДМФ) спречава ДСС-индуковани колитис преко активирања Нрф2 сигналног пута који је укључен у транслокацију Нрф2 нуклеарне јединице и инхибицију склопа инфламасома НЛРП3 [80].
Серија експеримената коришћењем природних и синтетичких једињења такође је открила инхибиторни ефекат Нрф2 на активацију инфламасома НЛРП3. На пример, лечење епигалокатехин-3-галата (ЕГЦГ) код мишева лупус нефритиса показало је смањење бубрежне активације инфламасома НЛРП3 која је посредована Нрф2 сигналним путем [81]. Слично, цитрал (3,7-диметил-2,6-октадиенал), главно активно једињење у кинеском биљном леку Литсеа цубеба, инхибира активацију НЛРП3 инфламасома преко Нрф2 антиоксидантног сигналног пута у моделу убрзаног и тешког лупус нефритиса (АСЛН) [82]. Слично, биоханин је штитио од повреде јетре изазване ЛПС/ГалН активирањем Нрф2 пута и инхибирањем активације НЛРП3 инфламазома код мужјака БАЛБ/ц мишева [83]. Штавише, показало се да мангиферин појачано регулише експресију Нрф2 и ХО-1 на начин зависан од дозе и инхибира јетрени НЛРП3, АСЦ, каспаза-1, ИЛ-1 изазван ЛПС/Д-ГалН? и ТНФ-? израз [84].
Упркос негативној регулацији НЛРП3 од стране Нрф2, он такође активира функцију инфламасома НЛРП3 и АИМ2. Хаитао Вен и колеге су открили то, Нрф2 ?/? мишји макрофаги су показали дефектну активацију НЛРП3 и АИМ2 инфламасома, али не и НЛРЦ4 инфламасома [85]. Занимљиво је да ово запажање приказује непознате функције Нрф2 у контексту болести повезаних са упалом; стога је веома важно даље проучавати како би се открио механизам у коме Нрф2 активира функцију инфламасома пре него што је размотримо као терапеутску мету.
Сузбијање транскрипције проинфламаторних цитокина од стране Нрф2
Недавно истраживање засновано на хроматинском имунопреципитацији (ЦхИП)-сек и ЦхИП-кПЦР резултатима код мишјих макрофага је открило да се Нрф2 везује за регионе промотера про-инфламаторних цитокина као што су ИЛ-6 и ИЛ-1? и инхибира регрутацију РНК Пол ИИ. Као резултат, РНК Пол ИИ није у стању да обради транскрипциону активацију ИЛ-6 и ИЛ-1? што на крају доводи до инхибиције експресије гена. По први пут, група Масајукија Јамамота открила је нови механизам којим Нрф2 не само да трансактивира своје низводне гене преко АРЕ већ и потискује транскрипциону активацију специфичних гена са или без АРЕ кроз инхибицију регрутовања РНК Пол ИИ [50].
Преслушавање између Нрф2 и НФ-?Б путева
НФ-?Б је протеински комплекс одговоран за транскрипцију ДНК који се налази у скоро свим врстама животињских ћелија и укључен је у различите процесе као што су запаљење, апоптоза, имуни одговор, раст и развој ћелија. п65, Рел протеин из породице НФ-?Б, има домен трансактивације, док п50 нема и захтева хетеродимеризацију са Рел протеином да би активирао транскрипцију. Током оксидативног стреса, И?Б киназа (ИКК) се активира и изазива фосфорилацију И?Б, што резултира ослобађањем и нуклеарном транслокацијом НФ-?Б. НФ-?Б изазива транскрипцију про-инфламаторних медијатора као што су ИЛ-6, ТНФ-а, иНОС, ИЛ-1 и интрацелуларна адхезија ЦОКС-2.
Абнормална регулација НФ-?Б повезана је са реуматоидним артритисом, астмом, инфламаторном болешћу црева и гастритисом изазваним инфекцијом Хелицобацтер пилори [86]. Тренутно се сматра да активност НФ-кБ утиче на Кеапл/Нрф2/АРЕ сигнални пут углавном у три аспекта: прво, Кеап1 деградира ИКК? кроз убиквитинацију, чиме се инхибира активност НФ-?Б [87]. Друго, инфламаторни процес индукује инфламаторне медијаторе као што је ЦОКС2 који потиче од циклопентенонског простагландина 15д-ПГЈ2, јаког електрофила који реагује са Кеап1 и активира Нрф2, чиме започиње транскрипцију гена уз истовремену инхибицију НФ-кБ [58] [88] Слика 3 А, Б). Треће, НФ-?Б се може комбиновати са компетитивним Нрф2 транскрипционим коактиватором ЦБП [89], [90] (Слика 3 Ц, Д).
Слика 3 Преслушавање између Нрф2 и НФ-?Б путева. (А) Кеап1 усмерава ИКК на ЦУЛ3 посредовану убиквитинацију и деградацију протеазома што на крају доводи до инхибиције НФ-?Б фосфорилације и овај механизам такође функционише као компетитивно везивање Нрф2 и ИКК са Кеап1. (Б) Оксидативни стрес активира ИКК који фосфорилише НФ-?Б, што доводи до његове транслокације у језгро и активације проинфламаторних цитокина као што је ЦОКС-2. Крајњи производ ЦОКС-2 познат као 15д-ПГЈ2 делује као индуктор Нрф2 који на крају доводи до супресије оксидативног стреса. (Ц) Нрф2 се везује са својим транскрипционим кофактором ЦБП заједно са малим Маф-ом и другим транскрипционим машинама да би покренуо експресију гена вођену АРЕ. (Д) Када се НФ-?Б веже са ЦБП на компетитивни начин, он инхибира везивање ЦБП са Нрф2, што доводи до инхибиције трансактивације Нрф2.
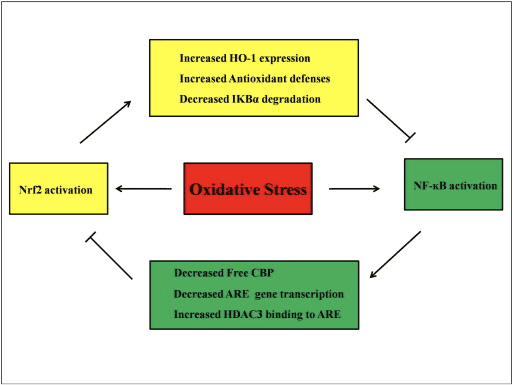
Претпоставља се да сигнални путеви Нрф2 и НФ-?Б међусобно делују како би контролисали транскрипцију или функцију низводних циљних протеина. Као оправдање ове претпоставке многи примери показују да се директна или индиректна активација и инхибиција дешавају између чланова Нрф2 и НФ-?Б путева (Сл. КСНУМКС). Као одговор на ЛПС, нокдаун Нрф2 значајно повећава активност транскрипције НФ-?Б и транскрипцију гена зависну од НФ-?Б, показујући да Нрф2 омета активност НФ-?Б [60], [91]. Поред тога, повећана експресија низводног ХО-2 зависног од Нрф1 инхибира активност НФ-аБ. Када су ћелије рака простате накратко изложене α-тохоферил сукцинату, деривату витамина Е, експресија ХО-1 се појачава. Крајњи производи ХО-1 инхибирају нуклеарну транслокацију НФ-?Б [92]. Ове ин виво студије сугеришу да Нрф2 негативно регулише НФ-кБ сигнални пут. ЛПС стимулише активност везивања НФ-?Б ДНК и ниво п65 подјединице НФ-?Б је значајно виши у нуклеарним екстрактима из плућа Нрф2?/? него од ВТ мишева, што указује на негативну улогу Нрф2 у активацији НФ-?Б. Штавише, Нрф2?/? фибробласти ембриона миша третирани ЛПС и ТНФ-? показују израженију активацију НФ-?Б узроковану ИКК активацијом и И?Б-? деградација [60]. А клиренс респираторног синцицијалног вируса је значајно смањен док је активност везивања НФ-?Б ДНК повећана у Нрф2?/? мишеви у поређењу са ВТ мишевима [93]. Пристаном изазван лупус нефритис у Нрф2?/? мишеви који су истовремено лечени сулфорафаном имају озбиљно оштећење бубрега и патолошке промене, као и повећану експресију иНОС и активацију НФ-?Б у поређењу са ВТ, што сугерише да Нрф2 побољшава лупус нефритис инхибирањем НФ-?Б сигналног пута и чишћењем РОС [94 ]. Активност НФ-аБ такође се јавља када се ћелије третирају са Нрф2 индуктором заједно са ЛПС и ТНФ-а. На пример, синтетички дериват халкона инхибира ТНФ-а-индуковану активацију НФ-пБ и директно и индиректно и делимично кроз индукцију експресије ХО-1 у хуманим интестиналним епителним ћелијама ХТ-29 [62]. Супресија НФ-?Б транслокације и активности везивања ДНК, као и супресија експресије иНОС у хепатоцитима, пронађена је када се Ф344 пацови лече 3Х-1,2-дитиол-3-тионом (Д3Т) [95]. Након заједничког третмана са сулфорафаном и ЛПС-ом, експресија иНОС, ЦОКС-2 и ТНФ-? у сировим 264.7 макрофага је смањена, што сугерише да сулфорафан има антиинфламаторну активност путем инхибиције везивања НФ-?Б ДНК [96]. Иако је урађено неколико експерименталних студија како би се објаснила веза између Нрф2 и НФ-?Б путева, остају супротстављени резултати. И позитивне и негативне регулације су пријављене између Нрф2 и НФ-кБ [97]. Обично, хемопревентивни електрофили 3Х-1,2-дитиол-3-тион, сулфорафан и тритерпеноид ЦДДО-Ме активирају Нрф2 инхибирањем НФ-кБ и његових смањених гена [98], [99], [100]. Насупрот томе, показало се да неколико агенаса или стања као што су РОС, ЛПС, напон смицања протока, оксидовани ЛДЛ и дим цигарета повећавају активност и Нрф2 и НФ-кБ [97]. Поред тога, ин виво студије су откриле да је активност НФ-кБ смањена у јетрима изолованим од Нрф2?/? мишева и активност везивања НФ-?Б је нижа у Нрф2?/? него код Нрф2+/+ мишева [101]. Међутим, ћелије ендотела људске аорте третиране аденовирусним вектором Нрф2 инхибирају низводне гене НФ-?Б без утицаја на активност НФ-?Б [8].
Слика 4 Регулаторна петља за Нрф2 и НФ-?Б. Нрф2 пут инхибира активацију НФ-?Б спречавањем деградације И?Б-? и повећање експресије ХО-1 и антиоксидативне одбране која неутралише РОС и хемикалије за детоксикацију. Као резултат, активација НФ-аБ повезана са РОС је потиснута. Слично, транскрипција посредована НФ-?Б смањује активацију Нрф2 смањењем�СУТранскрипција гена и слободни ЦРЕБ везујући протеин такмичећи се са Нрф2 за ЦБП. Штавише, НФ-?Б повећава регрутовање хистон деацетилазе (ХДАЦ3) у АРЕ регион и стога је спречена активација транскрипције Нрф2.
Активација Нрф2 сигналног пута игра главну улогу у експресији ензима и гена укључених у детоксикацију реактивних оксиданата тако што повећава антиоксидативни капацитет ћелија у људском телу. Иако су данас доступне многе истраживачке студије, регулаторни механизми у активацији Нрф2 нису у потпуности схваћени. Такође је пронађена могућа улога Нрф2 сигналног пута у лечењу упале. Др Алек Јименез ДЦ, ЦЦСТ Инсигхт
Улога Нрф2 у инфламаторним болестима
Ин виво студије су показале да Нрф2 игра важну улогу у инфламаторним болестима које утичу на различите системе; то укључује гастритис, колитис, артритис, пнеумонију, оштећење јетре, кардиоваскуларне болести, неуродегенеративне болести и оштећења мозга. У овим студијама, Нрф2?/? животиње су показале теже симптоме упале и оштећења ткива него ВТ животиње. Због тога се верује да сигнални пут Нрф2 има заштитни ефекат код инфламаторних болести. Интратрахеална уградња свињске панкреасне еластазе изазива хроничну опструктивну болест плућа, посебно емфизем. Мишеви са недостатком Нрф2 су веома подложни емфизему, а смањена експресија ХО-1, ПркИ и антипротеазног гена СЛПИ се јавља у алвеоларним макрофагима. Сматра се да је Нрф2 кључни регулатор у одбрамбеном систему посредованом макрофагом против повреда плућа [102]. Мишеви са недостатком Нрф2 са емфиземом изазваним излагањем дуванском диму током 6 месеци показују повећану бронхоалвеоларну упалу, појачану експресију маркера оксидативног стреса у алвеолама и повећану апоптозу алвеоларних септалних ћелија, што сугерише да Нрф2 делује против дуванске експресије антиоксида изазване дуваном. гени [102], [103]. Са поремећајем Нрф2, запаљење дисајних путева посредовано алергеном и астма коришћењем комплекса овалбумина показују повећану упалу дисајних путева, хиперреактивност дисајних путева, хиперплазију пехарастих ћелија и високе нивое Тх2 у бронхоалвеоларној лаважи и спленоцитима, док сигнални пут дисајних путева посредован Нрф2 ограничава сигналне путеве дисајних путева. , хиперсекреција слузи и хиперреактивност дисајних путева, као и индуковање многих антиоксидативних гена који спречавају развој астме [104]. Ињекција карагенана у плеуралну шупљину изазива плеуритис, а акумулација 15д-ПГЈ2 у Нрф2 инфламаторним ћелијама је ограничена на перитонеалне макрофаге миша. Током ране фазе упале, 15д-ПГЈ2 активира Нрф2 и регулише инфламаторни процес путем индукције ХО-1 и ПркИ. Студија је такође сугерисала да ЦОКС-2 има антиинфламаторни ефекат у раној фази производњом 15д-ПГЈ2 [105]. Орална примена 1% декстран сулфата натријум током 1 недеље изазива колитис повезан са хистолошким променама које укључују скраћивање крипта и инфилтрацију инфламаторних ћелија у ткиву дебелог црева. Да би заштитио интегритет црева код колитиса, Нрф2 би могао да игра важну улогу регулацијом проинфламаторних цитокина и индукцијом ензима за детоксикацију фазе ИИ [51]. У мишјем моделу плућне сепсе изазване ЛПС-ом, активност НФ-аБ регулише утицај инфламаторних цитокина као што су ЦОКС-2, ИЛ-2, ИЛ-113 и ТНФ? који су неопходни за покретање и подстицање упале [60]. Нрф2 смањује инфламаторно оштећење регулацијом ових инфламаторних фактора. У овим моделима акутне упале, повећана регулација антиоксидативних ензима, проинфламаторних цитокина и медијатора путем Нрф2 сигналног пута смањује инфламаторну повреду код ВТ животиња. Занимљиво је да је ово такође пријављено код Нрф2-нокаут мишева код којих су симптоми значајно погоршани у поређењу са ВТ мишевима.
Истраживање антиинфламаторних лекова зависних од Нрф2
Укратко, разговарали смо о експериментима који показују да пут сигнала Нрф2 игра регулаторну улогу у многим областима упале, тако да су антиинфламаторни агенси зависни од Нрф2 важни за лечење инфламаторних болести.
Биљке су биле изузетно богати извори једињења која активирају фактор транскрипције Нрф2, што доводи до појачане регулације цитопротективних гена. Недавно је спроведено неколико студија за испитивање деловања различитих антиинфламаторних агенаса, углавном биљног порекла. На пример, куркумин је активни састојак куркуме и такође се налази у малим количинама у ђумбиру; изотиоцијанати, посебно фенилизотиоцијанати су из броколија, целера и другог поврћа; а антоцијанини су из бобица и грожђа [124]. Истраживања су показала да сви ови агенси нису само добри антиоксиданти, већ имају и моћне антиинфламаторне ефекте путем Нрф2 индукције [125], [126]. Стога је развој нових антиинфламаторних Нрф2 активатора из биљног екстракта привукао велико интересовање у медицинским истраживањима.
Последњих година спроведени су многи експерименти на животињама да би се потврдило деловање ових једињења. Артесунате се углавном користи за тешку маларију, церебралну маларију и реуматске аутоимуне болести; ефикасан је и код септичких повреда плућа. Артесунат активира експресију Нрф2 и ХО-1, а потоњи смањује прилив проинфламаторних цитокина и леукоцита у ткиво да би спречио запаљење [127]. Сматра се да изовитексин, екстрахован из љуске пиринча Ориза сатива, има антиинфламаторна и антиоксидативна својства; игра заштитну улогу против акутне повреде плућа изазване ЛПС-ом тако што активира Нрф2/ХО-1 пут и инхибира МАПК и НФ-?Б [128]. Фимасартан, нови популарни блокатор рецептора ангиотензина ИИ који делује на систем ренин-ангиотензин, смањује крвни притисак; коришћење фимасартана за лечење мишева са хируршки изазваном једностраном опструкцијом уретера смањује оксидативни стрес, упалу и фиброзу преко регулације Нрф2 и антиоксидативног пута и инхибиције РАС и МАПК [129]. Сапанон је широко распрострањен у југоисточној Азији, где се користи као лек против грипа, антиалергијских и неуропротективних лекова; активира Нрф2 и инхибира НФ-?Б и тако може бити од користи у лечењу болести повезаних са Нрф2- и/или НФ-?Б [130]. Биксин екстрахован из семена Бикин ореллана се користи за инфективне и инфламаторне болести у Мексику и Јужној Америци; он смањује инфламаторне медијаторе, алвеоларно капиларно цурење и оксидативно оштећење на начин зависан од Нрф2 да би се ублажила повреда плућа изазвана вентилацијом и обновила нормална морфологија плућа [131]. Остала биљна једињења, као што су епигалокатехин галат, сулфорафан, ресвератрол, ликопен и екстракт зеленог чаја имају терапеутске ефекте на инфламаторне болести путем Нрф2 сигналног пута [132], [133], [134]. Недавно је пријављено да друга фитокемикалија, ериодиктиол, која је присутна у цитрусном воћу, има антиинфламаторне и антиоксидативне ефекте на повреду бубрега изазвану цисплатином и акутну повреду плућа изазвану сепсом регулацијом Нрф2, инхибирањем НФ-?Б и инхибицијом. експресија цитокина у макрофагима [135], [136]. Међутим, бројне фитокемикалије обећавају много за превенцију и лечење различитих људских болести, а неке су већ ушле у фазу клиничких испитивања (Табела 2).
Ова биљна једињења активирају Нрф2 сигнални пут углавном у облику електрофилних материјала који модификују цистеинске остатке Кеап1, што доводи до слободног нуклеарног Нрф2 везивања са АРЕ, што резултира активацијом транскрипције одговарајућег гена.
Сулфорафан и његови ефекти на рак, морталитет, старење, мозак и понашање, болести срца и друго
Изотиоцијанати су нека од најважнијих биљних једињења која можете добити у исхрани. У овом видеу радим за њих најсвеобухватнији случај који је икада направљен. Кратак распон пажње? Пређите на своју омиљену тему кликом на једну од временских тачака у наставку. Комплетна временска линија испод.
Кључни делови:
00:01:14 - Рак и смртност
00:19:04 - Старење
00:26:30 - Мозак и понашање
00:38:06 - Коначни приказ
00:40:27 - Доза
Комплетна временска линија:
00:00:34 - Увођење сулфорафана, главни фокус видео снимка.
00:01:14 - Потрошња крстастих поврћа и смањење смртности од свих узрока.
00:02:12 - Ризик од рака простате.
00:02:23 - Ризик од рака бешике.
00:02:34 - Рак плућа код пушача ризикује.
00:02:48 - Ризик од рака дојке.
00:03:13 - Хипотетично: шта ако већ имате рак? (интервенцијски)
00:03:35 – Уверљиви механизам који покреће асоцијативне податке о раку и морталитету.
00:04:38 - Сулфорафан и рак.
00:05:32 – Докази на животињама показују снажан ефекат екстракта клице броколија на развој тумора бешике код пацова.
00:06:06 - Ефекат директне суплементације сулфорафана код пацијената са раком простате.
00:07:09 - Биоакумулација метаболита изотиоцијаната у стварном ткиву дојке.
00:08:32 - Инхибиција матичних ћелија рака дојке.
00:08:53 - Лекција из историје: брассицас је утврдио да има здравствена својства чак и у старом Риму.
00:09:16 - Сулфорафанова способност да појача излучивање канцерогена (бензен, акролеин).
00:09:51 - НРФ2 као генетски прекидач преко елемената антиоксидативног одговора.
00:10:10 - Како активација НРФ2 појачава излучивање канцерогена преко глутатиона-С-коњугата.
00:10:34 - Кељ у Бриселу повећава глутатион-С-трансферазу и смањује оштећење ДНК.
00:11:20 - Напитак од клице брокуле повећава излучивање бензела за 61%.
00:15:45 - Конзумација крстастих поврћа и смртност од срчаних болести.
00:16:55 - Прашак клице брокуле побољшава липиде у крви и укупан ризик од болести срца код дијабетичара типа 2.
00:19:04 – Почетак секције старења.
00:19:21 – Исхрана обогаћена сулфорафаном продужава животни век буба од 15 до 30% (у одређеним условима).
00:20:34 - Значај слабе упале за дуговечност.
00:22:05 - Чини се да крстасто поврће и прах клица брокуле смањују широк спектар маркера упале код људи.
00:23:40 - Осврт на видео запис: рак, делови за старење
00:24:14 - Студије на мишевима сугеришу да сулфорафан може побољшати адаптивну имунолошку функцију у старости.
00:25:18 – Сулфорафан је побољшао раст косе на мишјем моделу ћелавости. Слика на 00:26:10.
00:26:30 - Почетак одељења за мозак и понашање.
00:27:18 - Ефекат екстракта клице брокуле на аутизам.
00:27:48 - Ефекат глукорафанина на шизофренију.
00:28:17 - Почетак дискусије о депресији (вероватни механизам и студије).
00:31:21 – Студија на мишу користећи 10 различитих модела депресије изазване стресом показује да је сулфорафан сличан ефикасан као флуоксетин (прозак).
00:32:00 - Студија показује да је директно уношење глукорафанина код мишева слично ефикасно у спречавању депресије из модела стреса социјалног пораза.
00:34:43 - Почетак одељења са трауматском повредом мозга.
00:35:01 - Сулфорафан убризган непосредно након ТБИ побољшава меморију (студија миша).
00:35:55 - Сулфорафан и неуронска пластичност.
00:36:32 – Сулфорафан побољшава учење у моделу дијабетеса типа ИИ код мишева.
00:37:19 – Сулфорафан и Дуцхенне мишићна дистрофија.
00:37:44 - Инхибиција миостатина у мишићним сателитским ћелијама (ин витро).
00:38:06 - Касни видео-приказ: смртност и рак, оштећење ДНК, оксидативни стрес и упале, излучивање бензена, кардиоваскуларне болести, дијабетес типа ИИ, ефекти на мозак (депресија, аутизам, шизофренија, неуродегенерација), пут НРФ2.
00:40:27 - Размишљања о проналажењу дозе клица брокуле или сулфорафана.
00:41:01 - Анегдоте о ницању код куће.
00:43:14 - О температурама кувања и активности сулфорафана.
00:43:45 - Конверзија бактерија у цревима сулфорафана из глукорафанина.
00:44:24 - Додаци делују боље у комбинацији са активном мирозиназом из поврћа.
00:44:56 - Технике кувања и крстасто поврће.
00:46:06 - Изотиоцијанати као гоитрогени.
Закључци
Тренутно се много истраживања фокусирало на улогу сигналног пута Нрф2/Кеап1/АРЕ у упали. Међу ензимима које појачава регулација Нрф2, ХО-1 је један од репрезентативних ензима одговора на стрес. ХО-1 има истакнута антиинфламаторна и антиоксидативна својства. Генерално, сигнални пут Нрф2 такође негативно регулише цитокине, факторе ослобађања хемокина, ММП и друге инфламаторне медијаторе ЦОКС-2 и иНОС производњу, који директно или индиректно утичу на релевантне НФ-кБ и МАПК путеве и друге мреже које контролишу упалу. Предлаже се да сигнални путеви Нрф2 и НФ-?Б међусобно делују како би регулисали транскрипцију или функцију низводних циљних протеина. Супресија или инактивација транскрипционе активности посредоване НФ-пБ преко Нрф2 највероватније се јавља у раној фази упале, пошто НФ-пБ регулише де ново синтезу низа проинфламаторних медијатора. Међутим, још увек постоје нека ограничења у истраживању као што су да ли постоје везе између Нрф2 и других сигналних путева као што је ЈАК/СТАТ, значај тренутних активатора Нрф2 добијених из природних биљних извора у запаљењу и како побољшати биолошку активност и побољшати циљање ових једињења. Ово захтева даљу експерименталну валидацију.
Поред тога, Нрф2 сигнални пут може регулисати > 600 гена [163], од којих > 200 кодира цитопротективне протеине [164] који су такође повезани са упалом, раком, неуродегенеративним болестима и другим великим болестима [165]. Све већи број доказа који сугеришу да је Нрф2 сигнални пут дерегулисан код многих карцинома, што доводи до аберантне експресије генске батерије зависне од Нрф2. Штавише, запаљење игра главну улогу у болестима повезаних са оксидативним стресом, посебно код рака. Примена неколико Нрф2 активатора за сузбијање упале може довести до аберантне експресије Нрф2 низводних гена који индукује онкогенезу и отпорност на хемотерапију и/или радиотерапију. Стога се могу развити високо специфични активатори Нрф2 да би се минимизирали његови плеиотропни ефекти. Неколико активатора Нрф2 показало је значајно побољшање антиинфламаторних функција код болести повезаних са оксидативним стресом. Најбољи пример активатора Нрф2 који је одобрила ФДА и који се широко користи за лечење инфламаторних болести као што је мултипла склероза (МС) је диметил фумарат. Тецфидера (регистровани назив диметил фумарата компаније Биоген) се ефикасно користи за лечење релапсирајућих облика мултипле склерозе код великог броја пацијената [152]. Међутим, ефикасност употребе активатора Нрф2 за лечење инфламаторних болести захтева даљу валидацију како би се избегли штетни ефекти Нрф2. Стога би развој терапија за антиинфламаторну активност посредовану Нрф2 могао имати значајан клинички утицај. Текуће студије сигналног пута Нрф2 широм света посвећене су развоју високо циљаних терапеутских агенаса за контролу симптома упале, као и за превенцију и лечење рака, као и неуродегенеративних и других великих болести.
У закључку, Нрф2 осећа нивое оксидативног стреса у људском телу и на крају помаже у промовисању регулације антиоксидативних и детоксикационих ензима и гена. Пошто је хронична упала узрокована повећаним нивоима оксидативног стреса повезана са неуродегенеративним болестима, Нрф2 може играти суштинску улогу у лечењу здравствених проблема попут Алцхајмерове болести, између осталих. Обим наших информација је ограничен на киропрактику и здравствене проблеме кичме. Да бисте разговарали о овој теми, слободно питајте др Хименеза или нас контактирајте на�915-850-0900 .
Додатна тема: Ублажавање болова у коленима без хируршке интервенције
Бол у колену је добро познат симптом који се може јавити услед разних повреда и/или стања колена, укључујући �спортске повреде. Колено је један од најсложенијих зглобова у људском телу јер се састоји од пресека четири кости, четири лигамента, разних тетива, два менискуса и хрскавице. Према Америчкој академији породичних лекара, најчешћи узроци болова у колену укључују сублуксацију пателе, пателарни тендинитис или скакачко колено и Осгоод-Сцхлаттерову болест. Иако се бол у колену највероватније јавља код људи старијих од 60 година, бол у колену се може јавити и код деце и адолесцената. Бол у колену се може лечити код куће према РИЦЕ методама, међутим, тешке повреде колена могу захтевати хитну медицинску помоћ, укључујући киропрактику. �
Неуродегенеративне болести, као што су Алцхајмерова болест и Паркинсонова болест, погађају милионе људи широм света. Доступне су различите опције лечења за лечење симптома неколико неуродегенеративних болести, иако су резултати често ограничени. Истраживања су показала да оксидативни стрес узрокован унутрашњим и спољашњим факторима може бити узрок развоја неуродегенеративних болести. Тхе фактор транскрипције, Нрф2, је утврђено да функционише као главни одбрамбени механизам против оксидативног стреса. Сврха чланка у наставку је да покаже ефекте Нрф2 о неуродегенеративним болестима.
Неуродегенеративне болести су повезане са акумулацијом специфичних протеинских агрегата, што указује на интимну везу између повређеног мозга и губитка протеостазе. Протеостаза се односи на све процесе помоћу којих ћелије контролишу обиље и савијање протеома захваљујући широкој мрежи која интегрише регулацију сигналних путева, експресију гена и системе деградације протеина. Овај преглед покушава да сумира најрелевантније налазе о транскрипционој модулацији протеостазе коју врши транскрипциони фактор НРФ2 (нуклеарни фактор (2-изведен из еритроида) сличан 2). НРФ2 се класично сматра главним регулатором антиоксидативног ћелијског одговора, иако се тренутно појављује као кључна компонента трансдукционе машинерије за одржавање протеостазе. Као што ћемо дискутовати, НРФ2 би се могао замислити као чвориште које компајлира хитне сигнале изведене из погрешно савијеног акумулације протеина како би се изградио координисан и трајан транскрипциони одговор. Ово се постиже функцијама НРФ2 које се односе на контролу гена укључених у одржавање физиологије ендоплазматског ретикулума, протеазома и аутофагије.
Нуклеарни фактор (2) налик 2 (НРФ2) је основни леуцин-зиппер протеин који се данас сматра главним регулатором ћелијске хомеостазе. Он контролише базалну експресију и експресију изазвану стресом преко 250 гена који деле заједнички појачивач цис деловања који се назива елемент антиоксидантног одговора (АРЕ) [1], [2], [3], [4], [5]. Ови гени учествују у реакцијама детоксикације фазе И, ИИ и ИИИ, метаболизму глутатиона и пероксиредоксина/тиоредоксина, производњи НАДПХ путем пентозофосфатног пута и јабучног ензима, оксидацији масних киселина, метаболизму гвожђа и протеостази [6]. С обзиром на ове широке цитопротективне функције, могуће је да један фармаколошки погодак у НРФ2 може ублажити ефекат главних криваца хроничних болести, укључујући оксидативни, инфламаторни и протеотоксични стрес. Улога НРФ2 у модулацији антиоксидативне одбране и решавању запаљења обрађена је у бројним студијама (прегледаним у [7]). Овде ћемо се фокусирати на његову улогу у протеостази, односно хомеостатској контроли синтезе протеина, савијања, трговине и деградације. Примери ће бити дати у контексту неуродегенеративних болести.
Губитак протеостазе утиче на активност НРФ2 код неуродегенеративних болести
Опште обележје неуродегенеративних болести је појава аберантне агрегације неких протеина. Дакле, погрешно савијени протеински агрегати ?-синуклеина (?-СИН) се налазе у Паркинсоновој болести (ПД), ?-амилоидним (А?) плаковима и хиперфосфорилисаним ТАУ неурофибриларним заплетима код Алцхајмерове болести (АД), Хунтингтина (Хтт) у Хантингтонова болест (ХД), супероксид дисмутаза 1 (СОД1) и ТАР ДНК везујући протеин 43 (ТДП-43) код амиотрофичне латералне склерозе (АЛС), прионски протеин (ПрП) код спонгиформних енцефалопатија, итд. Агрегати протеина могу имати утицај на неколико ћелијске путеве, што заузврат може утицати на нивое и активност НРФ2.
Различити слојеви регулације строго контролишу НРФ2 активност
У физиолошким условима, ћелије показују ниске нивое протеина НРФ2 због његовог брзог обртања. Као одговор на различите стимулусе, НРФ2 протеин се акумулира, улази у језгро и повећава транскрипцију гена који садрже АРЕ. Стога је управљање нивоима протеина НРФ2 кључна тачка која треба да интегрише позитивне и негативне улазне сигнале. Као што ћемо даље дискутовати, НРФ2 се активира различитим механизмима који се преклапају да би се оркестрирао брз и ефикасан одговор, али с друге стране, НРФ2 би могао бити инхибиран, вероватно у другој фази, како би се искључио његов одговор.
Са класичне тачке гледишта, активација НРФ2 се сматра последицом ћелијског одговора на оксидантна или електрофилна једињења. У том погледу, убиквитин Е3 лигазни адаптер Келцх-лике ЕЦХ-повезан протеин 1 (КЕАП1) игра кључну улогу. Молекуларни детаљи ће бити даље обрађени у одељку 4.1. Укратко, КЕАП1 делује као редокс сензор због критичних остатака цистеина који доводе до убиквитинације НРФ2 и протеазомске деградације. Поред ове класичне модулације, НРФ2 је дубоко регулисан сигналним догађајима. Заиста, показало се да различите киназе фосфорилишу и регулишу НРФ2. На пример, НРФ2 може бити фосфорилисан протеин киназама активираним митогеном (МАПК), иако његов допринос активности НРФ2 остаје нејасан [8], [9], [10], [11]. ПКА киназа као и неки ПКЦ изозими [12], ЦК2 [13] или Фин [14] фосфорилишу НРФ2 модификујући његову стабилност. Претходни рад наше групе је известио да гликоген синтаза кинс-3? (ГСК-3?) инхибира НРФ2 нуклеарним искључивањем и протеазомалном деградацијом [15], [25], [26], [27], [28], [29], [30]. О молекуларним детаљима ће се расправљати у одељку 4.1. Штавише, НРФ2 је подвргнут другим врстама прописа. На пример, ацетилација НРФ2 помоћу ЦБП/п300 повећава његову активност [17], док је инхибирају миР153, миР27а, миР142-5п и миР144 [16], или метилацијом острва цитозин-гуанин (ЦГ) унутар НРФ2 промотера. [18].
Утицај протеинских агрегата на регулаторне механизме НРФ2
У овом одељку ћемо се фокусирати на то како би акумулација погрешно савијеног протеина могла да утиче на активност НРФ2 дајући неке од путева поменутих горе као илустративне примере. Прво, морамо узети у обзир да је акумулација протеина уско повезана са оксидативним оштећењем. Заиста, погрешно савијена акумулација и агрегација протеина изазива абнормалну производњу реактивних врста кисеоника (РОС) из митохондрија и других извора [19]. Као што је горе поменуто, РОС ће модификовати цистеине КЕАП1 осетљиве на редокс, што доводи до ослобађања, стабилизације и нуклеарне локализације НРФ2.
Што се тиче протеинопатија, пример нерегулисаних сигналних догађаја који могу утицати на НРФ2 је дат хиперактивацијом ГСК-3? у огласу. ГСК-3?, такође познат као ТАУ киназа, учествује у фосфорилацији овог протеина повезаног са микротубулама, што резултира његовом агрегацијом, формирањем неурофибриларних сплетова и прекидом аксоналног транспорта (прегледано у [20]). С друге стране, ГСК-3? драматично смањује нивое и активност НРФ2 као што је горе поменуто. Иако није широко прихваћена, амилоидна каскада предлаже да токсични А? олигомери повећавају ГСК-3? активност заједно са ТАУ хиперфосфорилацијом и смрћу неурона [21], [22]. Постоје различити модели који објашњавају како А? фаворизује ГСК3-? активност. На пример, А? везује се за инсулински рецептор и инхибира ПИ3К и АКТ сигналне путеве, који су кључни за одржавање ГСК-3? инактивиран фосфорилацијом на свом Н-терминалном Сер9 остатку [23]. С друге стране, екстрацелуларни А? ступа у интеракцију са Фриззлед рецепторима, блокирајући ВНТ сигнализацију [24] и поново доводи до ослобађања активног ГСК-3?. Укратко, А? акумулација доводи до абнормалне хиперактивације ГСК-3?, чиме се нарушава одговарајући НРФ2 одговор.
Као што је објашњено у следећем одељку, погрешно савијени протеини доводе до активације ПЕРК и МАПК, који заузврат регулишу НРФ2 [31], [8], [9], [10], [11]. Штавише, дисрегулисана активност ЦБП/п300 је пријављена у неколико протеинопатија [32], а такође је приказано глобално смањење метилације ДНК у мозгу АД [33], што даје основу за истраживање релевантности ових налаза у регулацији НРФ2.
Ми и други смо на обдукцијама пацијената са ПД и АД приметили повећање нивоа НРФ2 протеина и неких његових мета, као што су хем оксигеназа 1 (ХМОКС1), НАДПХ кинон оксидаза 1 (НКО1), п62, итд., како имуноблотирањем тако и имунохистохемијом [34], [35], [36], [37], [38], [39]. Повећана регулација НРФ2 код ових болести тумачи се као неуспешан покушај оболелог мозга да поврати хомеостатске вредности. Међутим, друга студија је показала да је НРФ2 претежно локализован у цитоплазми хипокампалних неурона АД, што указује на смањену транскрипциону активност НРФ2 у мозгу [40]. Могуће је да је диспаритет ових запажања повезан са променама у факторима који контролишу НРФ2 дуж прогресивних фаза неуродегенерације.
Три главна система доприносе протеостази, а то су несавијени протеински одговор (УПР), убиквитин протеазомски систем (УПС) и аутофагија. Затим, представљамо доказе да замислимо НРФ2 као чвориште које повезује сигнале за хитне случајеве активиране протеинским агрегатима са машинама за деривате протеина.
НРФ2 учествује у Унфолдед Протеин Респонсе (УПР)
Активација НРФ2 као одговор на УПП
Оксидативно савијање протеина у ЕР је вођено бројним различитим путевима, од којих је најконзервативнији укључује протеин дисулфид-изомеразу (ПДИ) и сулфхидрил оксидазу ендоплазматски оксидоредуктин 1 (ЕРО1? и ЕРО1? код сисара) као дисулфидне донаторе. Укратко, ПДИ катализује формирање и кидање дисулфидних веза између остатака цистеина унутар протеина, док се они савијају, услед редукције и оксидације сопствених цистеинских аминокиселина. ПДИ се рециклира деловањем кућног ензима ЕРО1, који поново уводи дисулфидне везе у ПДИ [41]. Молекуларни кисеоник је терминални акцептор електрона ЕРО1, који генерише стехиометријске количине водоник пероксида за сваку произведену дисулфидну везу [42]. Пероксидазе (ПРКС4) и глутатион пероксидазе (ГПКС7 и ГПКС8) су кључни ензими за смањење водоник пероксида у ЕР. Када овај оксидо-редукциони систем не функционише како треба, долази до абнормалне акумулације погрешно савијених протеина у ЕР и скуп сигнала названих неодвијени протеински одговор (УПР) се преноси у цитоплазму и језгро да би се поново успоставила хомеостаза ЕР [43]. Идентификована су три протеина повезана са мембраном за откривање ЕР стреса код еукариота: активирајући фактор транскрипције 6 (АТФ6), ЕР еИФ2 панкреаса? киназа (ПЕРК, такође дволанчана РНК активирана протеин киназа слична ЕР киназа) и киназа 1 која захтева инозитол (ИРЕ1). Луминални домен сваког сензора је везан за шаперон од 78 кДа који се назива протеином регулисаним глукозом (ГРП78/БИП). БИП се дисоцира на ЕР стресу да би везао несавијене протеине, што доводи до активације три сензора [44].
НРФ2 и његов хомолог НРФ1, такође повезан са антиоксидативним одговором, учествују у трансдукцији УПР-а у језгро. У случају НРФ1, овај протеин се налази на ЕР мембрани и подлеже нуклеарној транслокацији након дегликозилације или цепања. Затим, активација УПР-а доводи до обраде НРФ1 и нуклеарне акумулације резултујућег фрагмента у нуклеарном одељку. Међутим, способност трансактивације гена који садрже АРЕ овог НРФ1 фрагмента је још увек у дискусији [45].
Гловер-Цуттер и сарадници су показали активацију НРФ2 ортолога Ц. елеганс, СКН-1, са различитим ЕР стресорима. Повећана експресија СКН-1 зависила је од различитих УПР медијатора, укључујући ИРЕ1 или ПЕРК ортологе црва [46]. У ћелијама са недостатком ПЕРК, поремећена синтеза протеина доводи до акумулације ендогених пероксида и касније апоптозе [47]. Ефектор који ПЕРК користи да заштити ЕР од ових пероксида може бити НРФ2, пошто је пријављено да ПЕРК фосфорилише НРФ2 на Сер40, спречавајући тако његову деградацију од стране КЕАП1 [31]. Индукција АСК1 ће такође вероватно играти улогу на овом путу кроз деловање киназе ИРЕ2 посредовано ТРАФ1 [48]. Иако је улога МАПК-а у регулацији НРФ2 и даље контроверзна, недавно је сугерисано да би пут ИРЕ1-ТРАФ2-АСК1-ЈНК могао активирати НРФ2 [49] (слика 1). Занимљиво је да у Ц. елеганс и људским ћелијама, нови докази сугеришу да цистеин сулфенилација ИРЕ1 киназе у њеној активационој петљи инхибира УПР посредован ИРЕ1 и покреће антиоксидативни одговор п38 покретан НРФ2. Подаци сугеришу да ИРЕ1 има древну функцију као цитоплазматски стражар који активира п38 и НРФ2 [50].
Слика 1. Уредба НРФ2 УПП. Акумулација несавијених или погрешно савијених протеина унутар ендоплазматског ретикулума може покренути одговор несавијеног протеина (УПР). Прво, цхаперон БИП се ослобађа из интралуминалног домена ЕР сензора ИРЕ1 и ПЕРК да би се везали несавијени / погрешно савијени протеини. Ово омогућава димеризацију и транс-ауто-фосфорилацију њихових цитосолних домена. ПЕРК активација резултира директном фосфорилацијом НРФ2 на Сер40, што доводи до транслокације НРФ2 у језгро и активације циљних гена. Активација ИРЕ1 индукује регрутовање ТРАФ2 праћено фосфорилацијом и активацијом АСК1 и ЈНК. Како је пријављено да ЈНК фосфорилише и активира НРФ2, разумно је мислити да би активација ИРЕ1 довела до повећане активности НРФ2.
Многе студије о индукцији УПР-а су спроведене са инхибитором гликозилације протеина туникамицином. Чини се да је НРФ2 неопходан за превенцију апоптотске смрти ћелија изазване туникамицином [31] и његова активација у овим условима је вођена аутофагном деградацијом КЕАП1 [51]. Сходно томе, схРНА посредовано утишавање експресије НРФ2 у ?ТЦ-6 ћелијама, мишји инсулином ?-ћелијској линији, значајно је повећало цитотоксичност изазвану туникомицином и довело до повећања експресије про-апоптотичког ЕР стресног маркера ЦХОП10. С друге стране, активација НРФ2 1,2-дитиол-3-тионом (Д3Т) смањила је цитотоксичност туникамицина и ослабила експресију ЦХОП10 и ПЕРК [52]. Занимљиво је да су олфакторни неурони подвргнути системској примени туникамицина повећали НРФ2 упоредо са другим УПР-члановима као што су ЦХОП, БИП, КСБП1 [53]. Ови резултати су проширени на студије ин виво, пошто је латерална вентрикуларна инфузија туникамицина код пацова изазвала експресију ПЕРК и НРФ2 у хипокампусу, праћену значајним когнитивним дефицитима, повећаном ТАУ фосфорилацијом и депозитима А?42 [54].
НРФ2 навише регулише кључне гене за одржавање физиологије ЕР
ЕР лумен треба обилно снабдевање ГСХ из цитосола да би се одржала хемија дисулфида. НРФ2 модулира кључне ензиме метаболизма ГСХ у мозгу, као што су транспорт цистин/глутамата, ?-глутамат цистеин синтетаза (?-ГС), каталитичке и модулаторске подјединице глутамат-цистеин лигазе (ГЦЛЦ и ГЦЛМ), глутатион редуктаза (ГР) глутатион пероксидаза (ГПКС) (прегледано у [55]). Релевантност НРФ2 у одржавању ГСХ у ЕР је подржана налазом да фармаколошка или генетска активација НРФ2 доводи до повећане синтезе ГСХ преко ГЦЛЦ/ГЦЛМ, док је инхибирање експресије ових ензима НРФ2-нокдауном изазвало акумулацију оштећених протеини унутар ЕР који доводе до активације УПР-а [56].
Код Ц. елеганс неколико компоненти УПР циљних гена регулисаних СКН-1, укључујући Ире1, Ксбп1 и Атф6. Иако НРФ2 појачано регулише експресију неколико гена пероксидазе (ПРКС) и глутатион пероксидазе (ГПКС) код сисара (прегледано у [57]), само је ГПКС8 веродостојни ЕР-локализован ензим, који садржи сигнал за проналажење КДЕЛ [58]. Губитак ГПКС8 изазива активацију УПР-а, цурење водоник пероксида добијеног из ЕРО1? у цитосол и ћелијску смрт. Водоник пероксид добијен из ЕРО1? активност не може да дифундује са ЕР у цитосол због усклађеног деловања ГПКС8 и ПРКС4 [59]. С тим у вези, анализа низа експресије гена за пут антиоксидативне одбране користећи РНК из дивљег типа и ткива мишева без НРФ2, открила је да је експресија ГПКС8 смањена у одсуству НРФ2 [60]. У складу са овим, анализа транскриптома узорака пацијената који пате од мијелопролиферативних неоплазми, полицитемије или мијелофиброзе, болести које су такође повезане са оксидативним стресом и хроничном инфламацијом ниског степена, показује ниже нивое експресије и НРФ2 и ГПКС8 у поређењу са контролним субјектима [61]. Још не постоје студије које посебно укључују ГПКС8 у заштиту људског мозга, али анализа транскриптома код мишева указује на компензаторно повећање ГПКС8 као одговор на Паркинсонов токсин МПТП [62].
Утицај НРФ2 на УПР дисрегулацију код неуродегенеративних болести
Неисправност ПДИ ензима и хронична активација УПР-а могу накнадно покренути или убрзати неуродегенерацију. Неурони захваћени болешћу, животињски модели неуродегенеративних болести, као и постмортем људска ткива доказали су повећање регулације неколико УПР-маркера у већини ових поремећаја. Промена ПДИ/УПР путање код неуродегенеративних болести је добро прегледана у [63], али треба узети у обзир следеће истакнуте тачке из узорака мозга након смрти. Нивои ПДИ су повећани у неуронима који носе замршеност и у Левијевим телима пацијената са АД и ПД [64], [65]. ПДИ и ЕРП57 су појачано регулисани у ЦСФ код пацијената са АЛС и у мозгу пацијената са ЦЈД [66], [67], [68]. БИП, ПЕРК, ИРЕ1 и АТФ6 су повишени у узорцима пацијената са АД, ПД или АЛС [69], [70], [71], [67]. БИП, ЦХОП и КСБП1 су повишени у постморталним узорцима мозга из ХД [72], [73]. Штавише, повећана регулација ЕРП57, ГРП94 и БИП пронађена је у ткивима кортекса код пацијената са ЦЈД [74]. Све у свему, ови докази откривају да акумулација погрешно савијених протеина у можданом паренхиму доводи до штетне и хроничне активације УПР-а. Занимљиво је да постоји недавна студија која повезује активацију НРФ2 од стране ПЕРК-а у раној АД. У овој студији, аутори су анализирали да ли промене посредоване оксидативним стресом у НРФ2 и УПР могу представљати ране догађаје у патогенези АД коришћењем људских ћелија периферне крви и АД трансгеног модела миша у различитим стадијумима болести. Повећан оксидативни стрес и повећани пСер40-НРФ2 примећени су у мононуклеарним ћелијама људске периферне крви изолованим од особа са благим когнитивним оштећењем. Штавише, они су пријавили поремећену хомеостазу ЕР калцијума и повећане маркере ЕР-стреса у овим ћелијама од особа са благим когнитивним оштећењем и благим АД [75].
Међусобна регулација НРФ2 и убиквитин протеазомског система (УПС)
УПС модулира нивое протеина НРФ2
УПС учествује у разградњи оштећених или погрешно савијених протеина и контролише нивое кључних регулаторних молекула у цитосолу и језгру. Централно језгро овог система је велики ензим са више подјединица који садржи протеолитички активни комплекс назван 20С. Протеазом језгра 20С разграђује несавијене протеине, али везивање за различите регулаторне протеинске комплексе мења његову специфичност и активност супстрата. На пример, додавање једне или две 19С регулаторне подјединице језгру 20С чини 26С протеазом и мења његову специфичност према природним савијеним протеинима [76], [77]. Протеазомска деградација захтева ковалентно везивање убиквитина. Коњугација убиквитина се одвија преко каскадног механизма у три корака. Прво, ензим Е1 који активира убиквитин активира убиквитин у реакцији која захтева АТП. Затим, један ензим Е2 (протеин носач убиквитин или ензим који коњугира убиквитин) преноси активирани убиквитин са Е1 на супстрат који је специфично везан за члана породице убиквитин-протеин лигазе, названог Е3. Иако ће тачна судбина убиквитинираног протеина зависити од природе ланца убиквитина, овај процес генерално доводи до разградње 26С протеазома [78].
Е3-лигаза КЕАП1 је најпознатији инхибитор НРФ2. Механизам регулације КЕАП1 елегантно објашњава како се нивои НРФ2 прилагођавају флуктуацијама оксиданса. У базалним условима, новосинтетизовани НРФ2 је захваћен хомодимером КЕАП1, који везује један НРФ2 молекул на две аминокиселинске секвенце са ниским (аспартат, леуцин, глицин; ДЛГ) и високим (глутамат, треонин, глицин, глутамат; ЕТГЕ). Интеракција са КЕАП1 помаже да се НРФ2 представи комплексу протеина ЦУЛЛИН3/РБКС1, што резултира његовом убиквитинацијом и накнадном протеазомалном деградацијом. Међутим, редокс модификација КЕАП1 омета представљање НРФ2 УПС-у који представља ЦУЛЛИН3/РБКС1. Као резултат, новосинтетизовани НРФ2 избегава деградацију зависну од КЕАП1, акумулира се у језгру и активира гене који садрже АРЕ [79], [80], [81], [82].
Адаптер Е3-лигазе ?-ТрЦП је такође хомодимер који учествује у сигналним догађајима везаним за фосфорилацију НРФ2 помоћу ГСК-3?. Ова киназа фосфорилише специфичне серинске остатке НРФ2 (аспартат, серин, глицин, изолеуцин серин; ДСГИС) да би се створио домен разградње који је затим препознат од ?-ТрЦП и означен за деградацију протеазома од стране ЦУЛЛИН1/РБКС1 комплекса. Идентификација специфичних амино киселина које су фосфорилисане помоћу ГСК-3? у овом дегрону је спроведена комбинацијом мутагенезе усмерене на место Нех6 домена, 2Д-гел електрофорезе [15], [26] и масене спектроскопије [83]. Сходно томе, инхибиција ГСК-3? високо селективним лековима или сиРНА против изоформа ГСК-3 резултирало је повећањем нивоа НРФ2 протеина. Слични резултати су пронађени са сиРНА против ?-ТрЦП изоформа 1 и 2. Стабилизација НРФ2 након ГСК-3? инхибиција се догодила у фибробластима мишјих ембриона са недостатком КЕАП1 и у ектопично експримираном НРФ2 делеционом мутанту коме недостају критични ЕТГЕ остаци за везивање високог афинитета за КЕАП1, што даље показује регулацију независну од КЕАП1.
У контексту неуродегенеративних болести, можемо замислити модулацију НРФ2 од стране УПС-а на два различита начина. С једне стране, КЕАП1 систем би осетио редокс неравнотежу насталу услед акумулације погрешно савијеног протеина, док би ГСК-3/?-ТрЦП оса деловала као активни учесник у сигналној трансдукцији измењеној губитком протеостазе (слика 2).
Слика 2 УПС чврсто контролише нивое НРФ2. У хомеостатским условима, ниски нивои НРФ2 се одржавају деловањем адаптера Е3 лигазе КЕАП1 и ?-ТрЦП. Лево, НРФ2 се везује за Келцх домене КЕАП1 хомодимера преко мотива ниског (ДЛГ) и високог (ЕТГЕ) афинитета. Преко свог БТБ домена, КЕАП1 се истовремено везује за ЦУЛЛИН3/РБКС1 комплекс, омогућавајући НРФ2 убиквитинацију и деградацију од стране 26 С протеазома. Штавише, ГСК-3? фосфорилише Сер335 и Сер338 остатке НРФ2 да би створио домен деградације (ДпСГИпСЛ) који је затим препознат од стране адаптера убиквитин лигазе ?-ТрЦП и означен за деградацију протеазома од стране ЦУЛЛИН3/РБКС1 комплекса. Десно, након излагања реактивним врстама кисеоника или електрофилима, критични Цис остаци у КЕАП1 су модификовани, чинећи КЕАП1 неспособним за ефикасну интеракцију са НРФ2 или ЦУЛЛИН3/РБКС1, а затим овај фактор транскрипције повећава свој полуживот и транскрипциону активност према АРЕ генима. Сигнални путеви који доводе до инхибиције ГСК-3?, као што је АКТ фосфорилација на Сер9, резултирају деградацијом НРФ2 оштећеном протеазомом, акумулацијом и индукцијом циљних гена.
НРФ2 повећава активност УПС-а кроз транскрипциону контролу протеазомских подјединица
НРФ2 регулише експресију неколико подјединица протеазома, штитећи тако ћелију од акумулације токсичних протеина. Чини се да је двадесет гена повезаних са протеазомом и убиквитинацијом регулисано НРФ2, према широкој анализи микромрежа из РНК јетре која је постављена са НРФ2 индуктором Д3Т [84]. У задњој студији, исти аутори су доказали да је експресија већине подјединица 26С протеазома повећана до три пута у јетри код мишева лечених Д3Т. Нивои протеина подјединице и активност протеазома су координирано повећани. Међутим, није примећена индукција код мишева код којих је транскрипциони фактор НРФ2 поремећен. Промоторска активност протеазомске подјединице ПСМБ5 (20С) се повећала или са прекомерном експресијом НРФ2 или са третманом активаторима у мишјим ембрионалним фибробластима, а АРЕ су идентификовани у проксималном промотеру ПСМБ5 [85]. Фармаколошка активација НРФ2 довела је до повишених нивоа експресије репрезентативних подјединица протеазома (ПСМА3, ПСМА6, ПСМБ1 и ПСМБ5) само у нестарећим људским фибробластима који садрже функционални НРФ2 [86]. Активација НРФ2 током адаптације на оксидативни стрес резултира високом експресијом ПСМБ1 (20С) и ПА28? подјединице (или С11, регулатор протеазома) [87]. Штавише, резултати из људских ембрионалних матичних ћелија открили су да НРФ2 контролише експресију протеина сазревања протеазома (ПОМП), протеазомског шаперона, који заузврат модулира пролиферацију самообнављајућих људских ембрионалних матичних ћелија, диференцијацију три заметна слоја и ћелијско репрограмирање [ 88]. Све заједно, ове студије показују да НРФ2 регулише експресију кључних компоненти УПС-а и стога активно доприноси чишћењу протеина који би иначе били токсични.
Ос НРФ2-УПС у неуродегенеративним болестима
Улога УПС-а у неуродегенеративним болестима је поље интензивне дебате. Почетне студије су пријавиле смањену активност протеазома у људским некропсијама пацијената погођених неколико неуродегенеративних болести. Међутим, друге студије које су користиле ин витро и ин виво приступе откриле су непромењену или чак повећану активност протеазома (прегледано у [89]). Једно могуће објашњење за ову неслагање је да се нивои компоненти УПС-а могу променити током прогресије болести иу различитим регионима мозга, као што је предложено за НРФ2-мете.
Упркос овој контроверзи, треба напоменути да ће регулација протеазомских гена који садрже АРЕ појачати УПС повећањем клиренса токсичних протеина у мозгу. Заиста, аблација НРФ1, такође модулатора антиоксидативног одговора, у неуронским ћелијама доводи до поремећене активности протеазома и неуродегенерације. Експерименти имунопреципитације хроматина и транскрипциона анализа показали су да је ПСМБ6 регулисан НРФ1. Поред тога, профилисање генске експресије довело је до идентификације НРФ1 као кључног регулатора транскрипције протеазомских гена у неуронима, што сугерише да пертурбације у НРФ1 могу допринети патогенези неуродегенеративних болести [90]. Занимљиво је да се показало да НРФ1 и његова дуга изоформа названа ТЦФ11 појачавају регулацију гена протеазома који садрже АРЕ након инхибиције протеасома у петљи повратне спреге како би се компензовала смањена протеолитичка активност [91], [92].
Што се тиче НРФ2, постоји корелација између смањења нивоа НРФ2, РПТ6 (19 С) и ПСМБ5 (20 С) у средњем мозгу мишева са недостатком ДЈ-1 који су третирани паракватом неуротоксина [93]. Штавише, природно једињење сулфорафан (СФН) даје робуснију слику НРФ2 као кључног модулатора УПС-а. Експерименти ин витро са Неуро2А ћелијама мишјег неуробластома показали су појачану експресију каталитичких подјединица протеазома, као и његове пептидазне активности као одговор на СФН. Овај лек је штитио ћелије од цитотоксичности посредоване водоник-пероксидом и оксидације протеина на начин који зависи од функције протеазома [94]. Поред тога, Лиу и сарадници су користили миша репортера да прате активност УПС-а као одговор на СФН у мозгу. Ови мишеви свеприсутно експримирају зелени флуоресцентни протеин (ГФП) спојен са сигналом конститутивне деградације који промовише његову брзу деградацију од стране УПС-а (ГФПу). У можданој кори, СФН је смањио ниво ГФПу са паралелним повећањем активности 5 С протеазома сличних химотрипсину (ПСМБ2), каспази (ПСМБ1) и трипсину (ПСМБ20). Поред тога, третман СФН ћелија добијених из Хунтингтона открио је да активација НРФ2 повећава деградацију мХтт и смањује цитотоксичност мХтт [95]. Главни механизам деловања СФН је кроз индукцију НРФ2 [96]. Специфичан допринос НРФ2 треба да се позабави употребом НРФ2-нулл система у даљим студијама.
Функционална веза између НРФ2 и макроаутофагије
Нивои НРФ2 протеина су модулисани помоћу адаптерског протеина П62
Аутофагија се односи на деградацију цитосолних компоненти унутар лизозома. Овај процес се користи за уклањање дуговечних и погрешно савијених протеина, као и оштећених органела. Директна веза између НРФ2 и аутофагије је први пут примећена у вези са адаптерским протеином п62, који се такође назива СКСТМ1 [97], [98], [99], [100], [101]. Овај протеин преноси убиквитиниране протеине до машина за протеазомску и лизозомску деградацију и секвестрира оштећене протеине у агрегате пре њихове деградације. П62 представља домен повезан са убиквитином (УБА), за везивање за убиквитиниране протеине, и регион интеракције са ЛЦ3 (ЛИР) за интеграцију са аутофагозомалном мембраном преко аутофагијског рецептора ЛЦ3.
Иако је п62 посредована индукција НРФ2 и његових циљних гена први пут објављена 2007. [102], молекуларни механизам није био у потпуности схваћен све до открића његове интеракције са КЕАП1 [103], [98], [99], [100] ], [101]. Коматсу и сарадници су идентификовали регион интеракције КЕАП1 (КИР) у п62 који је везивао КЕАП1 у истом основном површинском џепу као НРФ2 и са афинитетом везивања сличним ЕТГЕ мотиву у НРФ2, што сугерише конкуренцију између п62 и НРФ2. Показало се да фосфорилација Сер351 у КИР мотиву у п62 (349-ДПСТГЕ-354) повећава његов афинитет за КЕАП1, надмећући се са везивањем НРФ2 и омогућавајући његову акумулацију и транскрипциону активацију његових циљних гена [98], [99]. У ствари, прекомерна експресија п62 довела је до смањене убиквитинације НРФ2 и последичне стабилизације, као и индукције његових циљних гена [104]. Предложено је да неке киназе учествују у фосфорилацији п62. Циљ рапамицинског комплекса 1 (мТОРЦ1) код сисара може бити имплициран, пошто је третман са мТОР инхибитором рапамицином потиснуо фосфорилацију п62 и смањење КЕАП1 након третмана арсенитом. Недавно је показано да ТГФ-а-активирана киназа 1 (ТАК1) такође може да фосфорилише п62, побољшавајући деградацију КЕАП1 и регулацију НРФ2-уп-уп. Аутори ове студије сугеришу да је ово начин да се регулише ћелијска редокстаза у условима стабилног стања, пошто недостатак ТАК1 повећава РОС у одсуству било каквог егзогеног оксиданса у различитим ткивима миша, паралелно са смањењем нивоа протеина НРФ2 [105]. ].
Конструкт п62 коме недостаје УБА домен и даље је био способан да веже КЕАП1, што имплицира да интеракција не зависи од убиквитинираног КЕАП1 [101]. Међутим, хомолог п62 код Дросопхила меланогастер, назван Реф (2), не садржи КИР мотив и не ступа у директну интеракцију са ДмКЕАП1, иако се може везати за убиквитинирани ДмКЕАП1 преко УБА домена. Штавише, ДмКЕАП1 може директно да интерагује са Атг8 (хомолог ЛЦ3 сисара). Недостатак КЕАП1 доводи до индукције Атг8 и аутофагије у зависности од НРФ2 ортолога ЦнцЦ и независног од ТФЕБ/МИТФ [106]. Чини се да је однос између НРФ2 и аутофагије ипак очуван, наглашавајући његову функционалну релевантност.
Индукција НРФ2 од стране п62 је резултат и такмичења у везивању КЕАП1 и деградације КЕАП1 у лизозому. Утишавање п62 са сиРНА је удвостручило полуживот КЕАП1 упоредо са смањењем НРФ2 и његових циљних гена [101]. Сагласно, аблација експресије п62 је показала повећане нивое КЕАП1 у поређењу са мишевима дивљег типа. Веома релевантно, повећање нивоа КЕАП1 није било под утицајем инхибитора протеазома, али је смањено под аутофагијом која изазива гладовање [107]. У ствари, КЕАП1 је присутан у ћелијама сисара у аутофагним везикулама украшеним п62 и ЛЦ3 [99], [100], [103]. Сви ови подаци сугеришу да је КЕАП1 супстрат машинерије макроаутофагије, али ово питање треба детаљније анализирати због постојања неких контроверзних резултата. Нивои протеина КЕАП1 су повећани код Атг7-нулл мишева, кључног ефектора макроаутофагије [107], али фармаколошка инхибиција макроаутофагије са торин1, Е64/пепстатином или бафиломицином није успела да акумулира КЕАП1 [107], [100]. Све у свему, ови резултати сугеришу да повећани нивои п62 секвестрирају КЕАП1 у аутофагне вакуоле и вероватно ово резултира аутофагном деградацијом КЕАП1 омогућавајући активацију НРФ2 (слика 3). Две различите студије су известиле да редуктазе сулфинске киселине СЕСТРИНС играју важну улогу у овом контексту. СЕСТРИН 2 ступа у интеракцију са п62, КЕАП1 и РБКС1 и олакшава п62 зависну деградацију КЕАП1 и НРФ2 активације циљних гена [108]. Друга студија је показала да је СЕСТРИН 2 интераговао са УЛК1 и п62, промовишући фосфорилацију п62 на Сер403, што је олакшало деградацију теретних протеина укључујући КЕАП1 [109].
Слика 3 Нивои НРФ2 су регулисани адаптерским протеином п62. Фосфорилација Сер 351 у КИР мотиву п62 (349-ДПСТГЕ-354) помоћу мТОРЦ1, ТАК1 или других киназа резултира повећаним афинитетом за везивање за КЕАП1 због сличности са ЕТГЕ мотивом у НРФ2. Као последица тога, фосфориловани п62 замењује НРФ2 и везује КЕАП1. ЛИР мотив у п62 омогућава интеракцију са ЛЦ3 у аутофагозомалној мембрани, тако да се п62-КЕАП1 комплекс на крају разграђује у лизозому. Као последица тога, НРФ2 је у стању да се акумулира, транслоцира у језгро и повећа транскрипцију гена који садрже АРЕ, укључујући п62. Овај регулаторни механизам обезбеђује трајни одговор НРФ2, пошто КЕАП1 мора да се ново синтетише да би се инхибирала активност НРФ2.
Модулација гена макроаутофагије помоћу НРФ2
НРФ2 регулише експресију релевантних гена за макроаутофагију, као и за УПР и УПС. Први докази потичу из студија у којима се показало да се експресија п62 индукује при излагању електрофилима, РОС и азот оксиду [110], [111], [112]. Механизам индукције је описан неколико година касније са налазом да п62 садржи функционални АРЕ у свом промотору гена [99]. У недавној студији, неколико других функционалних АРЕ је пронађено и потврђено након биоинформатичке анализе и ЦхИП тестова. Штавише, ембрионални фибробласти миша и кортикални неурони из Нрф2-нокаут мишева показали су смањену експресију п62, која се могла спасити помоћу лентивируса који експримира НРФ2. Слично, недостатак НРФ2 је смањио нивое п62 у повређеним неуронима хипокампуса мишева [36]. Стога је сугерисано да активација НРФ2 повећава нивое п62, што резултира деградацијом КЕАП1 и фаворизује даљу стабилизацију НРФ2 у петљи позитивне повратне информације. Овај неканонски механизам индукције НРФ2 захтева промене у експресији гена и може бити релевантан одговор на продужени ћелијски стрес.
Показало се да је протеин за препознавање терета НДП52 транскрипцијски регулисан НРФ2. НДП52 ради на сличан начин као п62, препознајући убиквитиниране протеине и интеракцију са ЛЦ3 преко ЛИР домена, тако да се терет разграђује у лизозомима. Пет наводних АРЕ пронађено је у секвенци ДНК промотора Ндп52. Три од њих су идентификована са различитим мутантним конструктима и ЦхИП тестовима као неопходним за транскрипцију Ндп2 посредовану НРФ52 [113]. Треба напоменути да су нивои Ндп52 мРНА смањени у хипокампусу Нрф2-нокаут мишева. Једна од ових секвенци је такође потврђена у независној студији као АРЕ регулисана НРФ2 [36].
Међутим, улога НРФ2 у модулацији аутофагије није ограничена на индукцију ова два протеина за препознавање терета. Да би стекли дубљи увид у улогу НРФ2 у модулацији додатних гена повезаних са аутофагијом, наша група је прегледала базу података за имунопреципитацију хроматина ЕНЦОДЕ за два протеина, МАФК и БАЦХ1, који везују АРЕ регулисане НРФ2. Користећи скрипту генерисану из ЈАСПАР-ове консензус АРЕ секвенце, идентификовали смо неколико наводних АРЕ у многим генима аутофагије. Дванаест од ових секвенци је потврђено као НРФ2 регулисани АРЕ у девет гена аутофагије, чија је експресија смањена у фибробластима мишјих ембриона Нрф2-нокаут мишева, али би се могла обновити помоћу лентивируса који експримира НРФ2. Наша студија је показала да НРФ2 активира експресију неких гена укључених у различите кораке аутофагијског процеса, укључујући иницијацију аутофагије (УЛК1), препознавање терета (п62 и НДП52), формирање аутофагосома (АТГ4Д, АТГ7 и ГАБАРАПЛ1), елонгацију (АТГ2Б и АТГ5). ), и клиренс аутолизозома (АТГ4Д). Сходно томе, ток аутофагије као одговор на водоник пероксид био је поремећен када је НРФ2 био одсутан [36].
Релевантност експресије гена макроаутофагије посредоване НРФ2 у неуродегенеративним поремећајима
Показало се да дефектна аутофагија игра важну улогу у неколико неуродегенеративних болести [114], а аблација аутофагије доводи до неуродегенерације код мишева [115], [116]. Атг7-нокаут мишеви су открили да недостатак аутофагије доводи до акумулације п62 у инклузијским телима позитивним на убиквитин. КЕАП1 је издвојен у овим инклузивним телима, што је довело до стабилизације НРФ2 и индукције циљних гена [103]. Важно је да је прекомерна акумулација п62 заједно са убиквитинираним протеинима идентификована код неуродегенеративних болести, укључујући АД, ПД и АЛС [117]. У ствари, неурони који изражавају високе нивое АПП или ТАУ пацијената са АД такође су експримирали п62 и нуклеарни НРФ2, што сугерише њихов покушај да разграђују интранеуронске агрегате путем аутофагије [36].
Недостатак НРФ2 погоршава агрегацију протеина у контексту АД. У ствари, повећани нивои фосфорилисаног и саркозил-нерастворног ТАУ-а су пронађени код Нрф2-нокаут мишева, иако се никаква разлика у активностима киназе или фосфатазе није могла детектовати у поређењу са позадином дивљег типа [113]. Важно је да је показано да се НДП52 ко-локализује са ТАУ у мишјим неуронима, а директна интеракција између фосфо-ТАУ и НДП52 је приказана експериментима ко-имунопреципитације и на мишевима и на узорцима АД, указујући на његову улогу у деградацији ТАУ. Занимљиво је да је утишавање НДП52, п62 или НРФ2 у неуронима довело до повећања фосфо-ТАУ [113], [118]. Штавише, повећани интранеуронски АПП агрегати су пронађени у хипокампусу АПП/ПС1?Е9 мишева када је НРФ2 био одсутан. Ово је било у корелацији са измењеним маркерима аутофагије, укључујући повећане односе фосфо-мТОР/мТОР и фосфо-п70С6к/п70С6к (што указује на инхибицију аутофагије), повећане нивое пре-катепсина Д и већи број мултивезикуларних тела [119]. Код мишева који ко-експримирају хумани АПП (В717И) и ТАУ (П301Л), недостатак НРФ2 довео је до повећања нивоа укупног и фосфо-ТАУ у нерастворној фракцији и повећаних интранеуронских АПП агрегата, заједно са смањеним нивоима неурона п62, НДП52, УЛК1, АТГ5 и ГАБАРАПЛ1. Ко-локализација између адаптерског протеина п62 и АПП или ТАУ смањена је у одсуству НРФ2 [36]. Све у свему, ови резултати наглашавају важност НРФ2 у неуронској аутофагији.
Различити фактори транскрипције делују координирано на модулацију протеостазе
У условима стабилног стања, протеостаза се контролише интеракцијама протеин-протеин и пост-транслационим модификацијама чиме се добија брз одговор. Међутим, ћелијска адаптација захтева регулацију транскрипције гена УПР, УПС и аутофагије. Узимајући у обзир да су нервне ћелије континуирано изложене токсичним увредама ниског степена, укључујући оксидативни и протеотоксични стрес, појачање протеостазе изазвано модулацијом транскрипције може помоћи у спречавању дегенерације мозга.
У случају УПР-а, активација сваког од три крака ће коначно резултирати индукцијом транскрипције одређених гена (прегледано у [43]). На пример, фрагмент изведен из АТФ6 (АТФ6ф) се везује за ЕР-стресс респонсе елементс (ЕРСЕ) и индукује експресију неколико гена, укључујући КСБПИ, БИП и ЦХОП. Поред тога, ПЕРК сигнализација доводи до активације фактора транскрипције АТФ4, који контролише експресију више гена повезаних са УПР-ом и неких других, укључујући НРФ2 циљне гене Хмок1 и п62. Коначно, активација ИРЕ1 резултира стварањем активног фактора транскрипције, спојеног КСБП1 (КСБП1), који контролише транскрипцију гена који кодирају протеине укључене у савијање протеина.
С друге стране, показало се да је НРФ1 неопходан за експресију протеазомалних гена у мозгу, пошто су Нрф1-нокаут мишеви показали смањену експресију гена који кодирају различите подјединице 20С језгра, као и 19С регулаторни комплекс заједно са поремећеном протеазомалном функцијом [90 ]. И НРФ1 и НРФ2 се везују за АРЕ секвенце у промоторским регионима својих циљних гена, што сугерише да имају преклапајуће транскрипционе активности, иако се разликују по својим регулаторним механизмима и ћелијској локализацији [120].
Транскрипциони фактори породице Форкхеад бок О (ФОКСО) контролишу експресију више гена повезаних са аутофагијом. Слично ономе што се дешава са НРФ2, постоји више слојева регулације активности чланова ФОКСО, што може бити изазвано нутритивним или оксидативним стресом [121]. Коначно, фактор транскрипције ТФЕБ, који се сматра главним регулатором лизозомалне биогенезе, игра кључну улогу у регулацији аутофагије у условима нутритивног стреса. Дакле, инхибиција мТОРЦ1 доводи до нуклеарне транслокације ТФЕБ и индукције експресије гена аутофагије [122].
Све у свему, постојање различитих транскрипционих регулатора ових машина такође сугерише преслушавање и делимично редундантне механизме који могу да обезбеде протеостазу под различитим околностима. Сходно томе, НРФ2 може имати значајну улогу у ткивима која подржавају висок ниво оксидативног стреса. На пример, НРФ2 изазван оксидативним стресом може функционисати у условима богатим хранљивим материјама да би транскрипционо регулисао аутофагију, слично ономе што је пронађено за ТФЕБ у условима гладовања. Штавише, мозак углавном функционише у условима богатим хранљивим материјама, постављајући НРФ2 као релевантан механизам за активирање аутофагије у неуронима.
Обећавајући �терапеутски потенцијал за НРФ2 у протеинопатијама
У протеклих неколико година, направљен је велики напредак у познавању регулаторних улога УПР-а, УПС-а и аутофагије на активност НРФ2, као и реципрочне транскрипције компоненти ова три система посредоване НРФ2. Стога се могу појавити нове терапеутске могућности засноване на искоришћавању НРФ2 као кључног регулатора клиренса протеина код неуродегенеративних болести.
Међутим, кључно преостало питање је да ли ће бити корисно или штетно повећати нивое НРФ2 у мозгу. Анализа епидемиолошких података може дати делимичан одговор, јер указује на то да је ген НФЕ2Л2 високо полиморфан и да неки појединачни нуклеотидни полиморфизми који се налазе у његовом регулаторном региону промотера могу да обезбеде низ „физиолошких“ варијабилности у експресији гена на нивоу популације и неким хаплотиповима. били повезани са смањеним ризиком и/или одложеним почетком АД, ПД или АЛС [123]. Штавише, како су дискутовали Хаиес и колеге [124], ефекат НРФ2 може имати одговор у облику слова У, што значи да прениски нивои НРФ2 могу довести до губитка цитопротекције и повећане осјетљивости на стресоре, док превише НРФ2 може пореметити хомеостатску равнотежу према редуктивни сценарио (редукциони стрес), који би фаворизовао погрешно савијање и агрегацију протеина. Низак ниво НРФ2 у мозгу подржава идеју да би благо повећање регулације могло бити довољно за постизање користи у патолошким условима. У ствари, заштитна улога фармаколошке НРФ2 посредоване активације клиренса протеина је приказана у различитим културама ћелија неуродегенерације и ин виво моделима.
СФН је фармаколошки активатор НРФ2 за који је показано да индукује експресију гена протеазома и аутофагије [95], [36]. Занимљиво је да су Јо и његове колеге показали да СФН смањује нивое фосфорилисаног ТАУ-а и повећава Бецлин-1 и ЛЦ3-ИИ, што сугерише да активација НРФ2 може олакшати разградњу овог токсичног протеина путем аутофагије [113]. Штавише, деградација мХтт је побољшана са СФН, а ово је поништено употребом МГ132, што указује на протеазомалну деградацију овог токсичног протеина [95]. Пријављена је деградација фосфо- и нерастворљивог ТАУ-а посредована аутофагијом са органским флавоноидом физетином. Ово једињење је било у стању да изазове аутофагију истовремено промовишући активацију и нуклеарну транслокацију и ТФЕБ и НРФ2, заједно са неким од његових циљних гена. Овај одговор је спречен утишавањем ТФЕБ или НРФ2 [125]. Ботт и колеге су известили о корисним ефектима истовремених активатора НРФ2, НРФ1 и ХСФ1 на токсичност протеина код спиналне и булбарне мишићне атрофије, неуродегенеративног поремећаја узрокованог експанзијом ЦАГ понављања који кодирају полиглутамин у којима су присутни протеински агрегати [126]. Потенцијал активације НРФ2 за лечење неуродегенеративних поремећаја је демонстриран уз одобрење БГ-12, оралне формулације диметилфумарата индуктора НРФ2 (ДМФ), за лечење мултипле склерозе [127], [128]. Успех ДМФ-а код аутоимуних болести са јаком инфламаторном компонентом сугерише да би неуродегенеративне болести могле имати користи од репозиционирања овог лека. У недавној претклиничкој студији ?-синуклеинопатијског модела ПД, показало се да је ДМФ неуропротективан због, делимично, индукције аутофагије [129]. Студије које извештавају о корисним ефектима НРФ2 на неуродегенерацију, али се не фокусирају на његов ефекат на клиренс протеина, још су у изобиљу (за свеобухватан преглед, видети [7]). Ово је прилично релевантно, јер наглашава вишеструке штетне процесе који могу бити истовремено циљани једним ударцем у НРФ2, укључујући оксидативни стрес, неуроинфламацију или митохондријалну дисфункцију. Међутим, биће потребан будући рад да се дефинитивно утврди да ли фармаколошка активација НРФ2 може бити валидна стратегија за олакшавање разградње токсичних протеина у мозгу.
Као што је раније објашњено, погоршао ГСК-3? пријављена је активност код неуродегенеративних болести и спекулисало се да последична редукција НРФ2 може бити делимично одговорна за штетан исход. У овим патолошким условима, инхибитори ГСК-3 би такође могли да сарађују на повећању нивоа НРФ2 и протеостазе. Повољни ефекти инхибитора ГСК-3 су пријављени у различитим моделима неуродегенерације и, што је још интересантније, показало се да репресија ГСК-3 смањује нивое токсичних протеина [130], [131], [132], [133]. Иако још увек нису примећене директне везе између инхибиције ГСК-3 и НРФ2-транскрипционе регулације гена који промовишу протеостазу, разумно је спекулисати да би смањење активности ГСК-3 довело до повећања нивоа НРФ2, што ће на крају довести до појачаног протеостаза.
Транскрипциона активност НРФ2, као и ћелијски капацитет за одржавање протеостазе опадају са годинама, што је главни фактор ризика за развој неуродегенеративних болести. Разумно је мислити да би појачање НРФ2 и, последично, протеостазе, барем одложило акумулацију протеинских агрегата и неуродегенерацију. Заиста, третман људских сенесцентних фибробласта са тритерпеноидом 18а-глициретинске киселине (18а-ГА) је промовисао активацију НРФ2, што је довело до индукције протеазома и продуженог животног века. Ова студија сугерише да је фармаколошка активација НРФ2 могућа чак иу касној животној доби [86]. Штавише, каснија студија је показала да је ово једињење посредовало СКН-1 и активацији протеасома код Ц.елеганса са повољним ефектима на напредовање АД код релевантних модела нематода [134].
Узимајући све у обзир, чини се да је индукција гена повезаних са протеостазом посредована НРФ2 корисна у различитим протеинопатијама.
Сулфорафан и његови ефекти на рак, морталитет, старење, мозак и понашање, болести срца и друго
Изотиоцијанати су нека од најважнијих биљних једињења која можете добити у исхрани. У овом видеу радим за њих најсвеобухватнији случај који је икада направљен. Кратак распон пажње? Пређите на своју омиљену тему кликом на једну од временских тачака у наставку. Комплетна временска линија испод.
Кључни делови:
00:01:14 - Рак и смртност
00:19:04 - Старење
00:26:30 - Мозак и понашање
00:38:06 - Коначни приказ
00:40:27 - Доза
Комплетна временска линија:
00:00:34 - Увођење сулфорафана, главни фокус видео снимка.
00:01:14 - Потрошња крстастих поврћа и смањење смртности од свих узрока.
00:02:12 - Ризик од рака простате.
00:02:23 - Ризик од рака бешике.
00:02:34 - Рак плућа код пушача ризикује.
00:02:48 - Ризик од рака дојке.
00:03:13 - Хипотетично: шта ако већ имате рак? (интервенцијски)
00:03:35 – Уверљиви механизам који покреће асоцијативне податке о раку и морталитету.
00:04:38 - Сулфорафан и рак.
00:05:32 – Докази на животињама показују снажан ефекат екстракта клице броколија на развој тумора бешике код пацова.
00:06:06 - Ефекат директне суплементације сулфорафана код пацијената са раком простате.
00:07:09 - Биоакумулација метаболита изотиоцијаната у стварном ткиву дојке.
00:08:32 - Инхибиција матичних ћелија рака дојке.
00:08:53 - Лекција из историје: брассицас је утврдио да има здравствена својства чак и у старом Риму.
00:09:16 - Сулфорафанова способност да појача излучивање канцерогена (бензен, акролеин).
00:09:51 - НРФ2 као генетски прекидач преко елемената антиоксидативног одговора.
00:10:10 - Како активација НРФ2 појачава излучивање канцерогена преко глутатиона-С-коњугата.
00:10:34 - Кељ у Бриселу повећава глутатион-С-трансферазу и смањује оштећење ДНК.
00:11:20 - Напитак од клице брокуле повећава излучивање бензела за 61%.
00:15:45 - Конзумација крстастих поврћа и смртност од срчаних болести.
00:16:55 - Прашак клице брокуле побољшава липиде у крви и укупан ризик од болести срца код дијабетичара типа 2.
00:19:04 – Почетак секције старења.
00:19:21 – Исхрана обогаћена сулфорафаном продужава животни век буба од 15 до 30% (у одређеним условима).
00:20:34 - Значај слабе упале за дуговечност.
00:22:05 - Чини се да крстасто поврће и прах клица брокуле смањују широк спектар маркера упале код људи.
00:23:40 - Осврт на видео запис: рак, делови за старење
00:24:14 - Студије на мишевима сугеришу да сулфорафан може побољшати адаптивну имунолошку функцију у старости.
00:25:18 – Сулфорафан је побољшао раст косе на мишјем моделу ћелавости. Слика на 00:26:10.
00:26:30 - Почетак одељења за мозак и понашање.
00:27:18 - Ефекат екстракта клице брокуле на аутизам.
00:27:48 - Ефекат глукорафанина на шизофренију.
00:28:17 - Почетак дискусије о депресији (вероватни механизам и студије).
00:31:21 – Студија на мишу користећи 10 различитих модела депресије изазване стресом показује да је сулфорафан сличан ефикасан као флуоксетин (прозак).
00:32:00 - Студија показује да је директно уношење глукорафанина код мишева слично ефикасно у спречавању депресије из модела стреса социјалног пораза.
00:34:43 - Почетак одељења са трауматском повредом мозга.
00:35:01 - Сулфорафан убризган непосредно након ТБИ побољшава меморију (студија миша).
00:35:55 - Сулфорафан и неуронска пластичност.
00:36:32 – Сулфорафан побољшава учење у моделу дијабетеса типа ИИ код мишева.
00:37:19 – Сулфорафан и Дуцхенне мишићна дистрофија.
00:37:44 - Инхибиција миостатина у мишићним сателитским ћелијама (ин витро).
00:38:06 - Касни видео-приказ: смртност и рак, оштећење ДНК, оксидативни стрес и упале, излучивање бензена, кардиоваскуларне болести, дијабетес типа ИИ, ефекти на мозак (депресија, аутизам, шизофренија, неуродегенерација), пут НРФ2.
00:40:27 - Размишљања о проналажењу дозе клица брокуле или сулфорафана.
00:41:01 - Анегдоте о ницању код куће.
00:43:14 - О температурама кувања и активности сулфорафана.
00:43:45 - Конверзија бактерија у цревима сулфорафана из глукорафанина.
00:44:24 - Додаци делују боље у комбинацији са активном мирозиназом из поврћа.
00:44:56 - Технике кувања и крстасто поврће.
00:46:06 - Изотиоцијанати као гоитрогени.
Фактор 2 повезан са нуклеарним фактором 2 (НФ-Е2) везан за еритроиде, иначе познат као Нрф2, је фактор транскрипције који регулише експресију разних антиоксидативних и детоксикационих ензима. Истраживања су такође показала његову улогу у контроли оксидативног стреса. Већина неуродегенеративних болести, као што су Алцхајмерова болест и Паркинсонова болест, карактеришу оксидативни стрес и хронична упала, уобичајене мете Нрф2 приступи третману. Др Алек Јименез ДЦ, ЦЦСТ Инсигхт
Завршне напомене
Транскрипциони фактор НРФ2 оркестрира протеостатски одговор сенсингом и модулацијом промена у УПР, УПС и аутофагији (слика 4). Сходно томе, показало се да недостатак НРФ2 погоршава протеинопатију, што сугерише да је НРФ2 неопходан за оптимални клиренс протеина. Све заједно, можемо спекулисати да би НРФ2 могао бити занимљив терапеутски циљ за протеинопатије.
Слика 4 НРФ2 као чвориште које повезује хитне сигнале изведене из протеотоксика са заштитним транскрипционим одговором. Акумулација несавијених/погрешно савијених протеина ће довести до активације одговора несавијеног протеина (УПР) у ЕР. Активација ПЕРК или МАПК може довести до индукције транскрипције Гпк8 резидентног у ЕР-у и неколико ензима који регулишу нивое ГСХ, што је критично да би се обезбедило правилно савијање протеина. Протеински агрегати инхибирају активност протеазома (УПС), вероватно избегавајући деградацију НРФ2. Показало се да НРФ2 специфично модулира транскрипцију гена Псма3, Псма6, Псмб1, Псмб5 и Помп. Неколико других подјединица је регулисано на начин који зависи од НРФ2 као одговор на Д3Т, вероватно повећавајући листу подјединица протеазома регулисаних НРФ2. Аутофагија је главни пут за разградњу протеинских агрегата. Аутофагија такође регулише НРФ2, повезујући овај пут деградације са НРФ2 транскрипционом индукцијом п62, Ндп52, Улк1, Атг2б, Атг4ц, Атг5, Атг7 и Габарапл1.
Према претходном чланку, док се симптоми неуродегенеративних болести могу лечити различитим опцијама лечења, истраживачке студије су показале да активација Нрф2 може бити користан приступ лечењу. Јер Нрф2 активатори циљају широке механизме болести, све неуродегенеративне болести могу имати користи од употребе фактора транскрипције Нрф2. Налази Нрф2 су револуционирали лечење неуродегенеративних болести. Обим наших информација је ограничен на киропрактику и здравствене проблеме кичме. Да бисте разговарали о теми, слободно питајте др Хименеза или нас контактирајте на�915-850-0900 .
Додатна тема: Ублажавање болова у коленима без хируршке интервенције
Бол у колену је добро познат симптом који се може јавити услед разних повреда и/или стања колена, укључујући �спортске повреде. Колено је један од најсложенијих зглобова у људском телу јер се састоји од пресека четири кости, четири лигамента, разних тетива, два менискуса и хрскавице. Према Америчкој академији породичних лекара, најчешћи узроци болова у колену укључују сублуксацију пателе, пателарни тендинитис или скакачко колено и Осгоод-Сцхлаттерову болест. Иако се бол у колену највероватније јавља код људи старијих од 60 година, бол у колену се може јавити и код деце и адолесцената. Бол у колену се може лечити код куће према РИЦЕ методама, међутим, тешке повреде колена могу захтевати хитну медицинску помоћ, укључујући киропрактику. �
Оксидативни стрес се описује као оштећење ћелија узроковано слободним радикалима или нестабилним молекулима, што на крају може утицати на здраву функцију. Људско тело ствара слободне радикале да неутралише бактерије и вирусе, међутим, спољни фактори, као што су кисеоник, загађење и зрачење, често такође могу произвести слободне радикале. Оксидативни стрес је повезан са бројним здравственим проблемима.
Оксидативни стрес и други стресори укључују унутрашње заштитне механизме који могу помоћи у регулисању антиоксидативног одговора људског тела. Нрф2 је протеин који осећа ниво оксидативног стреса и омогућава ћелијама да се заштите од унутрашњих и спољних фактора. Такође је показано да Нрф2 помаже у регулацији гена који су укључени у производњу антиоксидативних ензима и гена на стресни одговор. Сврха доњег чланка је да објасни ефекти Нрф2 код рака.
Апстрактан
Пут Кеап1-Нрф2 је главни регулатор цитопротективних одговора на оксидативни и електрофилни стрес. Иако сигнални путеви ћелија покренути транскрипционим фактором Нрф2 спречавају покретање и напредовање рака у нормалним и премалигним ткивима, у потпуно малигним ћелијама активност Нрф2 пружа предност у расту повећавајући хеморезистенцију на карцином и појачавајући раст туморских ћелија. У овом графичком прегледу пружамо преглед пута Кеап1-Нрф2 и његове дисрегулације у ћелијама карцинома. Такође укратко сумирамо последице конститутивне активације Нрф2 у ћелијама карцинома и како се то може искористити у генској терапији карцинома.
Кључне речи:Нрф2, Кеап1, Рак, Антиоксидативни елемент одговора, Генска терапија
увод
Кеап1-Нрф2 пут је главни регулатор цитопротективних одговора на ендогене и егзогене стресове изазване реактивним врстама кисеоника (РОС) и електрофилима [1]. Кључни сигнални протеини унутар пута су фактор транскрипције Нрф2 (фактор 2 повезан са нуклеарним фактором еритроида 2) који се везује заједно са малим Маф протеинима за елемент антиоксидативног одговора (АРЕ) у регулаторним регионима циљних гена, и Кеап1 (Келцх ЕЦХ асоцирајући протеин 1), репресорски протеин који се везује за Нрф2 и промовише његову деградацију путем убиквитин протеазомског пута (слика 1). Кеап1 је протеин веома богат цистеином, мишји Кеап1 има укупно 25 и људски 27 цистеинских остатака, од којих већина може бити модификована ин витро различитим оксидантима и електрофилима [2]. Показало се да три од ових остатака, Ц151, Ц273 и Ц288, играју функционалну улогу мењајући конформацију Кеап1 што доводи до нуклеарне транслокације Нрф2 и накнадне експресије циљног гена [3] (слика 1). Тачан механизам у коме модификације цистеина у Кеап1 доводе до активације Нрф2 није познат, али два преовлађујућа, али не и међусобно искључива модела су (1) модел 'шарке и засуна', у којем Кеап1 модификације у остацима тиола који се налазе у ИВР-у Кеап1 поремете интеракцију са Нрф2 изазивајући неусклађеност лизинских остатака унутар Нрф2 који се више не могу полиубиквитиниловати и (2) модел у којем модификација тиола изазива дисоцијацију Цул3 од Кеап1 [3]. У оба модела, Кеап2 модификован индуктором и Нрф1 везан је инактивиран и, последично, новосинтетисани Нрф2 протеини заобилазе Кеап1 и транслоцирају се у језгро, везују се за АРЕ и покрећу експресију Нрф2 циљних гена као што је НАД(П)Х хинон оксидоредуктаза 1 (НКО1), хем оксигеназа 1 (ХМОКС1), глутамат-цистеин лигаза (ГЦЛ) и глутатион С трансферазе (ГСТ) (слика 2). Поред модификација Кеап1 тиола које резултирају индукцијом циљног гена Нрф2, протеини као што су п21 и п62 могу да се вежу за Нрф2 или Кеап1 и на тај начин ометају интеракцију између Нрф2 и Кеап1 [1], [3] (слика 3).
Слика 1. Структуре Нрф2 и Кеап1 и цистеински код. (А) Нрф2 се састоји од 589 аминокиселина и има шест еволуцијски високо конзервираних домена, Нех1-6. Нех1 садржи бЗип мотив, структуру основног региона - леуцинског затварача (Л-Зип), где је основни регион одговоран за препознавање ДНК, а Л-Зип посредује у димеризацији са малим Маф протеинима. Нех6 функционише као дегрон да посредује у деградацији Нрф2 у језгру. Нех4 и 5 су трансактивациони домени. Нех2 садржи ЕТГЕ и ДЛГ мотиве, који су потребни за интеракцију са Кеап1, и хидрофилни регион лизинских остатака (7 К), који су неопходни за Кеап1 зависну полиубиквитинацију и деградацију Нрф2. (Б) Кеап1 се састоји од 624 аминокиселинских остатака и има пет домена. Два мотива интеракције протеин-протеин, БТБ домен и Келцх домен, раздвојени су интервенционим регионом (ИВР). БТБ домен заједно са Н-терминалним делом ИВР посредује у хомодимеризацији Кеап1 и везивању са Цуллин3 (Цул3). Келцх домен и Ц-терминални регион посредују у интеракцији са Нех2. (Ц) Нрф2 реагује са два молекула Кеап1 кроз своје Нех2 ЕТГЕ и ДЛГ мотиве. И ЕТГЕ и ДЛГ се везују за слична места на доњој површини Кеап1 Келцх мотива. (Д) Кеап1 је богат цистеинским остацима, са 27 цистеина у хуманом протеину. Неки од ових цистеина налазе се у близини основних остатака и стога су одличне мете електрофила и оксиданата. Образац модификације остатака цистеина од стране електрофила је познат као цистеински код. Хипотеза цистеинског кода предлаже да структурно различита средства за активирање Нрф2 утичу на различите Кеап1 цистеине. Модификације цистеина доводе до конформационих промена у Кеап1, нарушавајући интеракцију између Нрф2 ДЛГ и Кеап1 Келцх домена, чиме се инхибира полиубиквитинација Нрф2. Показан је функционални значај Цис151, Цис273 и Цис288, јер су Цис273 и Цис288 потребни за супресију Нрф2 и Цис151 за активацију Нрф2 индукторима [1], [3].
Слика 2. Сигнални пут Нрф2-Кеап1. (А и Б) у базалним условима, два молекула Кеап1 везују се за Нрф2 и Нрф2 је полиубикуитилатед комплексом Е3 лигазе на бази Цул3. Ова полиубиквилилација резултира брзом разградњом Нрф2 од стране протеасома. Мали проценат Нрф2 излази из инхибиторног комплекса и акумулира се у језгру да посредује базалну експресију гена зависну од АРЕ, одржавајући тако ћелијску хомеостазу. (Ц) Под стресним условима, индуктори модификују цистеине Кеап1 што доводи до инхибиције свеприсутности Нрф2 дисоцијацијом инхибиторног комплекса. (Д) Према моделу шарки и засуна, модификација специфичних остатака цистеина Кеап1 доводи до конформационих промена у Кеап1 што резултира одвајањем Нрф2 ДЛГ мотива из Кеап1. Убиквитинација Нрф2 је поремећена, али везивање са ЕТГЕ мотивом остаје. (Е) У моделу дисоцијације Кеап1-Цул3, везивање Кеап1 и Цул3 је поремећено као одговор на електрофиле, што доводи до бекства Нрф2 из система убиквитинације. У оба предложена модела, индуктивно модификовани и за Нрф2 везани Кеап1 је инактивиран и, према томе, ново синтетисани Нрф2 протеини заобилазе Кеап1 и транслоцирају се у језгро, везују се за елемент антиоксидативног одговора (АРЕ) и подстичу експресију циља Нрф2 гени као што су НКО1, ХМОКС1, ГЦЛ и ГСТ [1], [3].
Слика 3. Механизми за конститутивну нуклеарну акумулацију Нрф2 у раку. (А) Соматске мутације у Нрф2 или Кеап1 нарушавају интеракцију ова два протеина. У Нрф2 мутације утичу на ЕТГЕ и ДЛГ мотиве, али у Кеап1 мутације су равномерније распоређене. Даље, онкогенска активација, као што је КрасГ12Д [5], или поремећај супресора тумора, као што је ПТЕН [11], може довести до индукције транскрипције Нрф2 и повећања нуклеарног Нрф2. (Б) Хиперметилација Кеап1 промотора у раку плућа и простате доводи до смањења експресије мРНК Кеап1, што повећава нуклеарну акумулацију Нрф2 [6], [7]. (Ц) У породичном папиларном бубрежном карциному, губитак активности ензима фумарат хидратазе доводи до акумулације фумарата и даље до сукцинације Кеап1 остатака цистеина (2СЦ). Ова пост-транслациона модификација доводи до прекида интеракције Кеап1-Нрф2 и нуклеарне акумулације Нрф2 [8], [9]. (Д) Акумулација протеина ометача као што су п62 и п21 може пореметити везивање Нрф2-Кеап1 и резултира повећањем нуклеарног Нрф2. п62 се везује за Кеап1 преклапајући џеп за везивање за Нрф2 и п21 директно комуницира са ДЛГ и ЕТГЕ мотивима Нрф2, надмећући се тако са Кеап1 [10].
Механизми активирања и дисрегулације Нрф2 у раку
Иако је цитопротекција обезбеђена активацијом Нрф2 важна за хемопревенцију рака у нормалним и премалигним ткивима, у потпуно малигним ћелијама активност Нрф2 пружа предност у расту повећавајући хеморезистенцију на карцином и појачавајући раст туморских ћелија [4]. Описано је неколико механизама помоћу којих се сигнални пут Нрф2 конститутивно активира код различитих карцинома: (1) соматске мутације у Кеап1 или Кеап1 везујућем домену Нрф2 нарушавајући њихову интеракцију; (2) епигенетско утишавање експресије Кеап1 што доводи до дефектне репресије Нрф2; (3) акумулација протеина ометача као што је п62 што доводи до дисоцијације комплекса Кеап1-Нрф2; (4) транскрипциона индукција Нрф2 онкогеним К-Рас, Б-Раф и ц-Миц; и (5) пост-транслациона модификација цистеина Кеап1 сукцинилацијом која се јавља у породичном папиларном карциному бубрега услед губитка активности ензима фумарат хидратазе [3], [4], [5], [6], [7], [ 8], [9], [10] (слика 3). Конститутивно обилни протеин Нрф2 узрокује повећану експресију гена који су укључени у метаболизам лекова, повећавајући тако отпорност на хемотерапијске лекове и радиотерапију. Поред тога, висок ниво протеина Нрф2 повезан је са лошом прогнозом рака [4]. Прекомерно активни Нрф2 такође утиче на пролиферацију ћелија усмеравањем глукозе и глутамина ка анаболичким путевима повећавајући синтезу пурина и утичући на пут пентоза фосфата да поспеши пролиферацију ћелија [11] (слика 4).
Слика 4. Двострука улога Нрф2 у туморегенези. У физиолошким условима, низак ниво нуклеарног Нрф2 довољан је за одржавање ћелијске хомеостазе. Нрф2 инхибира иницијацију тумора и метастазе карцинома елиминишући канцерогене, РОС и друге агенсе који оштећују ДНК. Током туморигенезе, гомилање оштећења ДНК доводи до конститутивне хиперактивности Нрф2, што помаже аутономним малигним ћелијама да издрже висок ниво ендогених РОС и да избегну апоптозу. Стално повишени нивои нуклеарног Нрф2 активирају метаболичке гене поред цитопротективних гена који доприносе репрограмирању метаболизма и појачаној пролиферацији ћелија. Карциноми са високим нивоом Нрф2 повезани су са лошом прогнозом због радио и хеморезистенције и агресивне пролиферације ћелија карцинома. Дакле, активност пута Нрф2 је заштитна у раним фазама туморигенезе, али штетна у каснијим фазама. Стога, за превенцију рака, појачавање активности Нрф2 остаје важан приступ, док је за лечење карцинома пожељна инхибиција Нрф2 [4], [11].
С обзиром да се висока активност Нрф2 обично јавља у ћелијама рака са негативним исходима, постоји потреба за терапијама за инхибицију Нрф2. Нажалост, због структурне сличности са неким другим члановима бЗип породице, развој специфичних инхибитора Нрф2 је изазован задатак и до данас је објављено само неколико студија о инхибицији Нрф2. Скринингом природних производа, Рен ет ал. [12] идентификовали су антинеопластично једињење брусатол као инхибитор Нрф2 који побољшава хемотерапеутску ефикасност цисплатина. Поред тога, инхибитори ПИ3К [11], [13] и Нрф2 сиРНА [14] су коришћени за инхибицију Нрф2 у ћелијама рака. Недавно смо користили алтернативни приступ, познат као генска терапија за самоубиство рака, да циљамо ћелије рака са високим нивоом Нрф2. Лентивирусни вектори вођени Нрф2 [15] који садрже тимидин киназу (ТК) се преносе у ћелије рака са високом АРЕ активношћу и ћелије се третирају пролеком, ганцикловиром (ГЦВ). ГЦВ се метаболише у ГЦВ-монофосфат, који се даље фосфорилише ћелијским киназама у токсични трифосфатни облик [16] (Слика 5). Ово доводи до ефикасног убијања не само туморских ћелија које садрже ТК, већ и суседних ћелија због ефекта посматрача [17]. АРЕ-регулисана ТК/ГЦВ генска терапија може се додатно побољшати комбиновањем хемотерапеутског агенса доксорубицина за рак са лечењем [16], подржавајући идеју да би овај приступ могао бити користан у комбинацији са традиционалним терапијама.
Слика 5. Суицидна генска терапија. Конститутивна акумулација нуклеарне киселине Нрф2 у ћелијама карцинома може се искористити употребом вирусног вектора покретаног Нрф2 за генску терапију самоубиства рака [16]. У овом приступу, лентивирусни вектор (ЛВ) који експримира тимидин киназу (ТК) под минималним промотором СВ40 са четири АРЕ трансдукује се у ћелије аденокарцинома плућа. Високи нуклеарни нивои Нрф2 доводе до снажне експресије ТК кроз везивање Нрф2. Ћелије се затим третирају пролеком, ганцикловиром (ГЦВ), који се фосфорилује помоћу ТК. Трифосфорилирани ГЦВ ремети синтезу ДНК и доводи до ефикасног убијања не само ТК које садрже туморске ћелије, већ и суседних ћелија због ефекта посматрача.
Нрф2 је главни регулатор који покреће производњу снажних антиоксиданата у људском телу који помажу у уклањању оксидативног стреса. Разни антиоксидативни ензими, попут супероксид дисмутазе или СОД, глутатион и каталазе, такође се активирају путем Нрф2. Даље, одређене фитокемикалије попут куркуме, ашваганде, бакопе, зеленог чаја и млека чичка активирају Нрф2. Истраживачке студије су то откриле Нрф2 активација може природно побољшати ћелијску заштиту и вратити равнотежу у људско тело.
Др Алек Јименез ДЦ, ЦЦСТ Инсигхт
Сулфорафан и његови ефекти на рак, морталитет, старење, мозак и понашање, болести срца и друго
Изотиоцијанати су нека од најважнијих биљних једињења која можете добити у исхрани. У овом видеу радим за њих најсвеобухватнији случај који је икада направљен. Кратак распон пажње? Пређите на своју омиљену тему кликом на једну од временских тачака у наставку. Комплетна временска линија испод.
Кључни делови:
00:01:14 - Рак и смртност
00:19:04 - Старење
00:26:30 - Мозак и понашање
00:38:06 - Коначни приказ
00:40:27 - Доза
Комплетна временска линија:
00:00:34 - Увођење сулфорафана, главни фокус видео снимка.
00:01:14 - Потрошња крстастих поврћа и смањење смртности од свих узрока.
00:02:12 - Ризик од рака простате.
00:02:23 - Ризик од рака бешике.
00:02:34 - Рак плућа код пушача ризикује.
00:02:48 - Ризик од рака дојке.
00:03:13 - Хипотетично: шта ако већ имате рак? (интервенцијски)
00:03:35 – Уверљиви механизам који покреће асоцијативне податке о раку и морталитету.
00:04:38 - Сулфорафан и рак.
00:05:32 – Докази на животињама показују снажан ефекат екстракта клице броколија на развој тумора бешике код пацова.
00:06:06 - Ефекат директне суплементације сулфорафана код пацијената са раком простате.
00:07:09 - Биоакумулација метаболита изотиоцијаната у стварном ткиву дојке.
00:08:32 - Инхибиција матичних ћелија рака дојке.
00:08:53 - Лекција из историје: брассицас је утврдио да има здравствена својства чак и у старом Риму.
00:09:16 - Сулфорафанова способност да појача излучивање канцерогена (бензен, акролеин).
00:09:51 - НРФ2 као генетски прекидач преко елемената антиоксидативног одговора.
00:10:10 - Како активација НРФ2 појачава излучивање канцерогена преко глутатиона-С-коњугата.
00:10:34 - Кељ у Бриселу повећава глутатион-С-трансферазу и смањује оштећење ДНК.
00:11:20 - Напитак од клице брокуле повећава излучивање бензела за 61%.
00:15:45 - Конзумација крстастих поврћа и смртност од срчаних болести.
00:16:55 - Прашак клице брокуле побољшава липиде у крви и укупан ризик од болести срца код дијабетичара типа 2.
00:19:04 – Почетак секције старења.
00:19:21 – Исхрана обогаћена сулфорафаном продужава животни век буба од 15 до 30% (у одређеним условима).
00:20:34 - Значај слабе упале за дуговечност.
00:22:05 - Чини се да крстасто поврће и прах клица брокуле смањују широк спектар маркера упале код људи.
00:23:40 - Осврт на видео запис: рак, делови за старење
00:24:14 - Студије на мишевима сугеришу да сулфорафан може побољшати адаптивну имунолошку функцију у старости.
00:25:18 – Сулфорафан је побољшао раст косе на мишјем моделу ћелавости. Слика на 00:26:10.
00:26:30 - Почетак одељења за мозак и понашање.
00:27:18 - Ефекат екстракта клице брокуле на аутизам.
00:27:48 - Ефекат глукорафанина на шизофренију.
00:28:17 - Почетак дискусије о депресији (вероватни механизам и студије).
00:31:21 – Студија на мишу користећи 10 различитих модела депресије изазване стресом показује да је сулфорафан сличан ефикасан као флуоксетин (прозак).
00:32:00 - Студија показује да је директно уношење глукорафанина код мишева слично ефикасно у спречавању депресије из модела стреса социјалног пораза.
00:34:43 - Почетак одељења са трауматском повредом мозга.
00:35:01 - Сулфорафан убризган непосредно након ТБИ побољшава меморију (студија миша).
00:35:55 - Сулфорафан и неуронска пластичност.
00:36:32 – Сулфорафан побољшава учење у моделу дијабетеса типа ИИ код мишева.
00:37:19 – Сулфорафан и Дуцхенне мишићна дистрофија.
00:37:44 - Инхибиција миостатина у мишићним сателитским ћелијама (ин витро).
00:38:06 - Касни видео-приказ: смртност и рак, оштећење ДНК, оксидативни стрес и упале, излучивање бензена, кардиоваскуларне болести, дијабетес типа ИИ, ефекти на мозак (депресија, аутизам, шизофренија, неуродегенерација), пут НРФ2.
00:40:27 - Размишљања о проналажењу дозе клица брокуле или сулфорафана.
00:41:01 - Анегдоте о ницању код куће.
00:43:14 - О температурама кувања и активности сулфорафана.
00:43:45 - Конверзија бактерија у цревима сулфорафана из глукорафанина.
00:44:24 - Додаци делују боље у комбинацији са активном мирозиназом из поврћа.
00:44:56 - Технике кувања и крстасто поврће.
00:46:06 - Изотиоцијанати као гоитрогени.
priznanja
Овај рад подржали су Финска академија, Фондација Сигрид Јуселиус и финске организације за борбу против рака.
У закључку, нуклеарни фактор (из еритроида 2) сличан 2, такође познат као НФЕ2Л2 или Нрф2, је протеин који повећава производњу антиоксиданата који штите људско тело од оксидативног стреса. Као што је горе описано, стимулација Нрф2 пута се проучава за лечење болести узрокованих оксидативним стресом, укључујући рак. Обим наших информација је ограничен на киропрактику и здравствене проблеме кичме. Да бисте разговарали о овој теми, слободно питајте др Хименеза или нас контактирајте на�915-850-0900 .
Додатна тема: Ублажавање болова у коленима без хируршке интервенције
Бол у колену је добро познат симптом који се може јавити услед разних повреда и/или стања колена, укључујући �спортске повреде. Колено је један од најсложенијих зглобова у људском телу, јер се састоји од пресека четири кости, четири лигамента, различитих тетива, два менискуса и хрскавице. Према Америчкој академији породичних лекара, најчешћи узроци болова у колену укључују сублуксацију пателе, тендинитис пателарног или колена скакача и Осгоод-Сцхлаттерову болест. Иако се болови у колену највероватније јављају код људи старијих од 60 година, болови у колену могу се јавити и код деце и адолесцената. Болови у колену могу се лечити код куће следећи РИЦЕ методе, међутим, тешке повреде колена могу захтевати хитну медицинску помоћ, укључујући хиропрактичку негу.
ДНК подржава приближно 20,000 гена, од којих сваки има програм за стварање протеина или ензима потребних за здрав начин живота. Сваки од ових образаца треба стално да се регулише неком врстом „промотора“ који управља тачно колико се сваке супстанце и/или хемикалије генерише и под којим условима ће се оне такође развијати.
Повезивањем са одређеном врстом промоторских подручја сличних прекидачу, познатим као елемент антиоксидативног одговора, или АРЕ, Нрф2 фактор�подржава брзину стварања стотина различитих гена који омогућавају ћелијама да преживе под стресним околностима. Ови гени затим стварају селекцију антиоксидативних ензима који развијају одбрамбену мрежу неутрализацијом оксиданата и чишћењем токсичних нуспроизвода који су остали у њиховој производњи, поред тога што помажу у обнављању оштећења које су изазвали.
Шта је оксидативни стрес?
Неколико оксиданата попут супероксидног радикала, или О2-., и водоник пероксида, или Х2О2, створено је праксом сагоревања супстанци и/или хемикалија које одржавају људско тело. Људско тело поседује антиоксидативне ензиме који неутралишу и детоксикују реактивну храну и пиће које конзумирамо. Нрф2 модулира њихову производњу како би одржао равнотежу и наглашава потражњу за свим овим ензимима. Ова равнотежа може бити прекинута због неколико фактора, укључујући године.
Како старимо, људско тело ствара мање Нрф2 и ова деликатна равнотежа може постепено почети да се окреће ка оксидативној страни, стању које се назива оксидативни стрес. Болест такође може изазвати прекомерну производњу оксиданата. Инфекције, алергије и аутоимуни поремећаји могу додатно покренути наше имуне ћелије да стварају реактивне оксиданте, као што је О2-. , Х2О2, ОХ и ХОЦл, где се здраве ћелије оштећују и реагују упалом. Болести повезане са старењем, укључујући срчани удар, мождани удар, рак и неуродегенеративна стања попут Алцхајмерове болести, такође повећавају развој оксиданата, стварајући стрес и одговор на упалу.
Шта су Нрф2 активатори?
Протеин Нрф2, такође назван фактор транскрипције због начина на који може да подржи и контролише ензиме и гене, је тајни елемент низа биохемијских реакција унутар ћелије који реагује на модификације у когнитивној равнотежи, као и на оксидативну равнотежу. Сензорни елементи овог пута модификују и испуштају Нрф2, покрећући га тако да се може проширити у језгро ћелије према ДНК. Нрф2 може алтернативно да укључи или искључи гене и ензиме које подржава да заштити ћелију.
На срећу, разне супстанце које су активатори Нрф2 развијају се кроз конзумацију одређених биљака и екстраката који су коришћени пре неколико векова у традиционалним лековима Кине и Индијанаца. Чини се да су ове фитокемикалије подједнако моћне са мање нуспојава као и фармацеутски производи који активирају Нрф2 који се данас користе.
Фактор везан за нуклеарни фактор еритроид 2, познатији као Нрф2, је фактор транскрипције који штити ћелију регулацијом гена, ензима и антиоксидативних одговора. Транскрипцијски фактори су врста протеина који се везује за ДНК како би промовисао стварање специфичних супстанци и хемикалија, укључујући глутатион С-трансферазе, или ГСТ. Активација Нрф2 индукује производњу активних протеина који показују моћан антиоксидативни капацитет који помажу у смањењу оксидативног стреса.
Др Алек Јименез ДЦ, ЦЦСТ Инсигхт
Наука иза активације Нрф2
Након што је 2. створен почетни додатак исхрани који активира Нрф2004, било је познато минималне информације о функцији Нрф2 пута. Приближно 200 новина у литератури о Нрф2, такође познатом као нуклеарни фактор сличан 2 или НФЕ2Л2, постојало је и истраживачи су тек почели да откривају антиоксидативни одговор Нрф2 код сисара. Међутим, од 2017. године штампано је преко 9,300 академских истраживачких студија о овом „главном регулатору“.
У стварности, Нрф2 регулише многе антиоксидативне ензиме који нису у корелацији са генима, већ нуде заштиту од разних околности повезаних са стресом са којима се сусрећу ћелије, органи и на крају организми, под здравим и патолошким условима. На основу ове нове количине информација из објављених академских истраживачких студија, истраживачи се сада могу боље развијати Нрф2 дијететски суплементи.
Од 2007. године, истраживачке студије су показале сложену функцију Нрф2 пута. Утврђено је да активатори Нрф2 опонашају факторе различитих структура унутар људског тела. Кроз ове путеве, Нрф2 активатори су опремљени да осете променљиве услове у целој ћелији како би одржали равнотежу и одговорили на еволуирајуће захтеве гена.
Зашто користити суплементе који активирају Нрф2?
Како се способности Нрф2-активације смањују са годинама у организмима, промене могу почети да се дешавају. Истраживачке студије су показале да фокус Нрф2 у ћелијама опада са годинама, показујући повећане маркере оксидативног стреса. Услед ових промена могу се развити различите болести повезане са узрастом, као што су атеросклероза и кардиоваскуларне болести, артритис, рак, гојазност, дијабетес типа 2, хипертензија, катаракта и Алцхајмерова болест, као и Паркинсонова болест. Оксидативни стрес је пронађен код ових здравствених проблема.
Стимулисањем капацитета ћелије да повећа производњу Нрф2 активатора, Нрф2 дијететски суплементи може помоћи да се оживи сопствена способност људског тела да се супротстави ефектима оксидативног стреса. Полинезасићене масне киселине, или ПУФА, су једни од молекула који се најлакше оксидују и посебно су подложни оштећењу слободних радикала. Производња тиобарбитурне киселине, или ТБАРС, може се повећати са годинама, што указује на повећан оксидативни стрес заједно са падом Нрф2 регулисаних путева.
Биолошки гледано, индукција гена је заиста спор механизам, који генерално захтева неколико сати за пренос кроз пут. Као резултат тога,�многи ензими поседују сопствене прекидаче за укључивање/искључивање који се могу покренути за неколико минута различитим регулаторним ензимима. Истраживачи су развили власничке композиције Нрф2 активатора који користе ову базу знања о активацији. Активација Нрф2 се састоји не само од тога што се фактор транскрипције Нрф2 ослобађа из свог инхибитора и мигрира у језгро ћелије, већ се такође везује за специфичне ДНК секвенце како би се подстакла експресија цитопротективног гена, регулишући темпо којим се Нрф2 уклања из језгра.
Разумевање поступка елиминације и активације Нрф2 у људском телу омогућило је истраживачима да изграде комбинације различитих активатора Нрф2 да би постигли рефлексију гена кроз његову модулацију. Комбинација базе знања, заједно са широким спектром других истраживачких студија, помогла је да се произведу Нрф2 активатори за употребу као дијететски суплементи. Обим наших информација је ограничен на киропрактику и здравствене проблеме кичме. Да бисте разговарали о овој теми, слободно питајте др Хименеза или нас контактирајте на�915-850-0900 .
Курирао др Алек Јименез
Додатна тема: Ублажавање болова у коленима без хируршке интервенције
Бол у колену је добро познат симптом који се може јавити услед разних повреда и/или стања колена, укључујући �спортске повреде. Колено је један од најсложенијих зглобова у људском телу, јер се састоји од пресека четири кости, четири лигамента, различитих тетива, два менискуса и хрскавице. Према Америчкој академији породичних лекара, најчешћи узроци болова у колену укључују сублуксацију пателе, тендинитис пателарног или колена скакача и Осгоод-Сцхлаттерову болест. Иако се болови у колену највероватније јављају код људи старијих од 60 година, болови у колену могу се јавити и код деце и адолесцената. Болови у колену могу се лечити код куће следећи РИЦЕ методе, међутим, тешке повреде колена могу захтевати хитну медицинску помоћ, укључујући хиропрактичку негу.
Антиоксиданси се научно називају једињењима која ограничавају процес оксидације у људском телу, а ако се не контролишу, могу створити слободне радикале који могу развити бројне ланчане реакције које могу изазвати оштећење ћелија. На срећу, људско тело може да створи такве уграђене имуне механизме, међутим, када се реактивне врсте кисеоника, или РОС, не могу неутралисати, замислите сићушни пламен који измиче контроли када се унесе кисеоником, штета ће се сигурно догодити. .
Да наставимо са ширењем метафоре пламена, крајњи производ неспособности да неутралише утицај РОС, или реактивних врста кисеоника, је оштећење као и запаљење, другим речима, људско тело је буквално у пламену. Фантастична ствар је да постоје антиоксиданси који могу изузетно помоћи у борби против овог здравственог проблема, а овај антиоксиданс је глутатион. Иако је откривен 1889. године, антиоксидативни ефекат глутатиона постао је једна од најзанимљивијих тема у савременим истраживањима.
Мајстор антиоксиданата: глутатион
Моћна супстанца је трипептид који се развија из цистеина, глутаминске киселине и глицина. Због своје способности да заштити људско тело од стварања слободних радикала, глутатион на крају може помоћи у промовисању здравог имунолошког система. На бази Научни извештаји у 2015, утврђено је да способност глутатиона да функционише синергистички са пероксиредином и каталазом помаже у заштити ћелија од водоник пероксида. Ова синергистичка формула функционише против реактивних врста кисеоника, или РОС. Глутатион, пероксидредин и каталаза су есенцијални елементи у повећању ћелијске хомеостазе, што је суштински процес здравих ћелија, ткива и органа у целини.
Поред тога, глутатион повећава укупну структуру и функцију имуног система користећи свој важан ефекат на функције лимфоцита. Према Катедра за имунохемију, правилно допуњавање нивоа глутатиона у људском телу може значајно побољшати имунолошке реакције. На пример, два рандомизована плацебо контролисана испитивања показала су да је терапијски третман имунокомпромитованих пацијената са Н-ацетил-цистеином, или НАЦ, резултирао, у оба случаја, у значајном порасту већине имунолошких процеса који су укључивали потпуно подмлађивање. активности природних ћелија убица. Н-ацетил-цистеин, или НАЦ, користи сумпор из глутатиона и комбинује га са отровним молекулима, који тада постају растворљиви у води и испуштају се у људско тело.
Глутатион такође има способност да ревитализује липоинску киселину, као и да рециклира витамине Ц и Е, који су неопходни да би се покренули одређени системски процеси слањем електрона да неутралишу слободне радикале. На основу истраживачке студије из ПЛоС ОНЕ, глутатионом погођени пацијенти са дијабетес метиллусом, или Т2ДМ, и микобактеријом туберкулозе. Нормално, особе са слабим имунолошким системом имају тенденцију да покажу већу изложеност М. тб, или микобактерија туберкулозе, болести или инфекције. Штавише, особе са дијабетесом метиллусом типа 2, или Т2ДМ, су два до три пута склоније туберкулози од људи без Т2ДМ. Истраживачка студија је такође сугерисала да је повећање нивоа глутатиона у макрофагима изолованим од пацијената са Т2ДМ довело до побољшане контроле М.Тб болести или инфекције. Ови резултати показују да нижи нивои глутатиона код пацијената са Т2ДМ доприносе повећању шансе за М. тб болест или инфекцију. Штавише, зависно од Диетро Гхеззи на медицинској школи у Брајтону и Сасексу, оксидативни стрес може на крају изазвати лошу структуру и функцију имуног система.
На срећу, глутатион игра кључну улогу у јачању и контроли имунитета. На пример, глутатион је неопходан за урођене и адаптивне процесе унутар имунолошког система, укључујући пролиферацију Т-лимфоцита, фагоцитну активност полиморфонуклеарних неутрофила и функције дендритских ћелија, које могу бити фундаменталне јер се састоје од ћелија које представљају антиген. . Ћелијски медитирани имунитет укључује протеинске антигене који у почетку почињу да дегенеришу у ендоцитним везикулама макрофага и дендритских ћелија, стога се мањи пептиди показују на површини да активирају пролиферацију антиген специфичних Т ћелија. Поред тога, глутатион помаже у стварању цитокина, а неопходно је одржавати производњу интерферона-гама дендритских ћелија, што је важно за заштиту од интрацелуларних патогена укључујући микобактерије.
Н-ацетил-цистеин, или НАЦ, који се научно назива прекурсор глутатиона, такође је веома моћан ћелијски антиоксиданс који се користи као антиоксидант за хватање слободних радикала. Уобичајено познат по својој улози у спречавање токсичности ацетаминофена, НАЦ, или �Н-ацетил-цистеин, доказано је да поседује неколико здравствених и добробити. Према Целл Јоурнал, НАЦ помаже у подршци здравом инфламаторном одговору и може позитивно утицати на термин и превремене порођаје код људи. Истраживачка студија је закључила да је код жена са претходним превременим порођајем и бактеријском вагинозом, 0.6 грама НАЦ дневно узиманих орално заједно са прогестероном након 16. недеље трудноће, штитило од рецидива превременог порођаја и побољшало неонатални исход. У закључку, откривени су и позитивни ефекти НАЦ-а на изградњу мишића. Након три минута упорних контракција, дошло је до 15 процената побољшаног учинка, што показује како НАЦ игра основну улогу у побољшању изградње мишића и смањењу укупног умора током порођаја.
Истраживачи су такође открили да НАЦ, или Н-ацетил-цистеин, може бити од користи онима који имају синдром полицистичних јајника или ПЦОС. ПЦОС, или синдром полицистичних јајника, је уобичајена болест ендокриних жлезда која погађа отприлике 5 до 10 процената жена у репродуктивном добу. Код таквих пацијената постоји већи ризик од метаболичког синдрома, где је употреба НАЦ-а помогла да се поврати здрав ниво инсулина и осетљивост.
Увид др Алек Јименез-а
Глутатион се назива „господаром антиоксиданата“ због његове фундаменталне улоге у постизању и одржавању општег здравља и добробити. Док је људско тело способно да производи сопствени глутатион, лоша исхрана, загађење, токсини, прекомерна употреба лекова и/или лекова, стрес, трауме, старење, болести и зрачење могу смањити наше природне нивое глутатиона. Ово заузврат може учинити појединце подложнијим оштећењу ћелија услед оксидативног стреса, слободних радикала, инфекција и рака. Суплементација глутатионом стога може имати огромне користи за људско тело. Заједно са алтернативним опцијама лечења, као што је киропрактика, нивои глутатиона се поново могу регулисати како би се побољшало благостање.
Поред тога, здравствени радници су предложили примену суплементације глутатионом заједно са другим алтернативним опцијама лечења, као што су нега киропрактике, за даље побољшање општег здравља и добробити. Антиоксиданси су важни за одржавање максималног благостања, као и за инхибирање ланчане реакције слободних радикала који узрокују оштећење или оштећење ћелија. Моћни антиоксиданси попут глутатиона, као што је претходно поменуто, на крају помажу у регулисању развоја ових слободних радикала и обезбеђују здравији одговор имуног система. Истраживачке студије су то откриле нега киропрактике може такође играти суштинску улогу у овом процесу, природно подстичући активност антиоксиданата у људском телу. Киропрактичка нега је безбедан и ефикасан приступ третману који користи прилагођавање кичме и ручне манипулације за исправљање кичмених неслагања, или сублуксација, како би се омогућило људском телу да се природно излечи без употребе лекова/лекова и/или хируршких интервенција.
Коначно, антиоксиданси показују своја биолошка својства кроз велики број здравствених користи, које би сада могле бити неопходне више него икада са сваким порастом навале стреса, болести и загађења у нашем савременом свету, што све доприноси оштећењу и/или оштећењу ћелија. . Глутатион и његов прекурсор, НАЦ, или Н-ацетил-цистеин, настављају да показују свој моћан статус у области антиоксиданата. Заједно са алтернативним опцијама лечења, као што је киропрактика, људи могу да искористе све предности које овај моћни антиоксиданс може да понуди. Обим наших информација је ограничен на киропрактику, као и на повреде и стања кичме. Да бисте разговарали о овој теми, слободно питајте др Хименеза или нас контактирајте на�915-850-0900 .
Курирао др Алек Јименез
Додатне теме: Бол у леђима
Бол у леђима је један од најчешћих узрока инвалидности и пропуштених дана на послу широм света. Заправо, болови у леђима приписани су као други најчешћи разлог посета лекарској ординацији, надмашени само инфекцијама горњих дисајних путева. Отприлике 80 процената становништва ће се барем једном током живота суочити са неком врстом болова у леђима. Кичма је сложена структура коју између осталих меких ткива чине кости, зглобови, лигаменти и мишићи. Због тога су повреде и / или отежани услови, као нпр хернија дискова, на крају може довести до симптома болова у леђима. Спортске повреде или повреде у аутомобилским несрећама често су најчешћи узрок болова у леђима, али понекад најједноставнији покрети могу имати болне резултате. Срећом, алтернативне опције лечења, попут хиропрактичке неге, могу помоћи у ублажавању болова у леђима употребом кичмених подешавања и ручних манипулација, на крају побољшавајући ублажавање болова.
Киропрактичар др Александар Хименез баци поглед оксидативни стрес, шта је то, како утиче на тело и антиоксидантну одбрану да поправи ситуацију.
др Есра Бирбен, 1 др Умит Мурат Сахинер, 1 др Кансин Сакесен, 1 др Серпил Ерзурум, 2 и Омер Калајци, 1
Апстракт: Реактивне врсте кисеоника (РОС) производе живи организми као резултат нормалног ћелијског метаболизма и фактора околине, као што су загађивачи ваздуха или дим цигарета. РОС су високо реактивни молекули и могу оштетити ћелијске структуре као што су угљени хидрати, нуклеинске киселине, липиди и протеини и променити њихове функције. Промена равнотеже између оксиданата и антиоксиданата у корист оксиданата назива се �оксидативни стрес.� Регулација редукционог и оксидационог (редок) стања је критична за виталност ћелије, активацију, пролиферацију и функцију органа. Аеробни организми имају интегрисане антиоксидативне системе, који укључују ензимске и неензимске антиоксиданте који су обично ефикасни у блокирању штетних ефеката РОС. Међутим, у патолошким условима, антиоксидативни системи могу бити преоптерећени. Оксидативни стрес доприноси многим патолошким стањима и болестима, укључујући рак, неуролошке поремећаје, атеросклерозу, хипертензију, исхемију/перфузију, дијабетес, синдром акутног респираторног дистреса, идиопатску плућну фиброзу, хроничну опструктивну болест плућа и астму. У овом прегледу, сумирамо ћелијске оксидативне и антиоксидативне системе и расправљамо о ћелијским ефектима и механизмима оксидативног стреса.
Кључне речи: антиоксиданс, оксиданс, оксидативни стрес, реактивне врсте кисеоника, редокс
(ВАО Јоурнал 2012; 5:9)
Реактивне врсте кисеоника (РОС) производе живи организми као резултат нормалног ћелијског метаболизма. При ниским до умереним концентрацијама, функционишу у физиолошким процесима ћелије, али при високим концентрацијама производе штетне модификације ћелијских компоненти, као што су липиди, протеини и ДНК.1�6 Промена равнотеже између оксиданса/антиоксиданса у корист оксиданата назива се �оксидативни стрес.� Оксидативни стрес доприноси многим патолошким стањима, укључујући рак, неуролошке поремећаје,7�10 атеросклерозу, хипертензију, исхемију/перфузију,11�14 дијабетес, синдром акутног респираторног дистреса, идиопатску плућну фиброзу, хроничну опструктивну болест плућа ,15 и астма.16�21 Аеробни организми имају интегрисане антиоксидативне системе,� који укључују ензимске и неензимске антиоксиданте који су обично ефикасни у блокирању штетних ефеката РОС. Међутим, у патолошким условима, антиоксидативни системи могу бити преоптерећени. У овом прегледу сумирамо ћелијске оксидативне и антиоксидативне системе и регулацију редукционог и оксидационог (редок) стања у здравственим и болесним стањима.
ОКСИДАНТС
Ендогени извори РОС
РОС се производе из молекуларног кисеоника као резултат нормалног ћелијског метаболизма. РОС се могу поделити у 2 групе: слободни радикали и нерадикали. Молекули који садрже један или више неспарених електрона и тиме дају реактивност молекулу називају се слободни радикали. Када 2 слободна радикала деле своје неспарене електроне, стварају се нерадикални облици. Три главна РОС која су од физиолошког значаја су супероксид ањон (О3.), хидроксилни радикал (ОХ) и водоник-пероксид (Х22О2). РОС су сажети у табели 2.
Супероксид ањон се формира додавањем 1 електрона молекуларном кисеонику.22 Овај процес је посредован никотин аденин динуклеотид фосфат [НАД(П)Х] оксидазом или ксантин оксидазом или митохондријалним системом за транспорт електрона. Главно место за производњу супероксид ањона су митохондрије, машинерија ћелије за производњу аденозин трифосфата. Обично се електрони преносе кроз митохондријски ланац транспорта електрона ради редукције кисеоника у воду, али отприлике 1 до 3% свих електрона цури из система и производи супероксид. НАД(П)Х оксидаза се налази у полиморфонуклеарним леукоцитима, моноцитима и макрофагима. Након фагоцитозе, ове ћелије производе експлозију супероксида који доводи до бактерицидне активности. Супероксид се претвара у водоник пероксид дејством супероксид дисмутаза (СОД, ЕЦ 1.15.1.1). Водоник пероксид лако дифундује кроз плазма мембрану. Водоник пероксид се такође производи ксантин оксидазом, аминокиселинском оксидазом и НАД(П)Х оксидазом�23,24 и у пероксизомима потрошњом молекуларног кисеоника у метаболичким реакцијама. У низу реакција које се називају Хабер-Вајсове и Фентонове реакције, Х2О2 се може разградити до ОХ2 у присуству трансмисионих метала попут Фе21 или Цу21.25
Фе31 +�.О2�?Фе2 +�О2 Хабер Веисс
Фе2 +�Х2О2�?Фе3 +�ОХ�+ .ОХ Фентонова реакција
О 2 такође може да реагује са Х2 О2 и генерише ОХ.26,27 Хидроксилни радикал је најреактивнији од РОС и може оштетити протеине, липиде, угљене хидрате и ДНК. Такође може да покрене пероксидацију липида узимањем електрона из полинезасићених масних киселина.
Гранулоцитни ензими даље проширују реактивност Х2О2 преко еозинофил пероксидазе и мијелопероксидазе (МПО). Код активираних неутрофила, Х2О2 троши МПО. У присуству хлоридног јона, Х2О2 се претвара у хипохлорну киселину (ХОЦл). ХОЦл је веома оксидативан и игра важну улогу у убијању патогена у дисајним путевима.28 Међутим, ХОЦл такође може да реагује са ДНК и индукује интеракције ДНК-протеина и производи производе оксидације пиримидина и додаје хлорид базама ДНК.29,30 Еозинофил пероксидаза и МПО такође доприносе оксидативном стресу модификацијом протеина халогенацијом, нитрацијом и умрежавањем протеина преко тирозил радикала.31�33
Други слободни радикали добијени кисеоником су пероксилни радикали (РОО$). Најједноставнији облик ових радикала је хидропероксилни радикал (ХОО$) и има улогу у пероксидацији масних киселина. Слободни радикали могу покренути ланчане реакције пероксидације липида апстраховањем атома водоника из метиленског угљеника бочног ланца. Липидни радикал затим реагује са кисеоником да би произвео пероксилни радикал. Пероксилни радикал покреће ланчану реакцију и трансформише полинезасићене масне киселине у липидне хидропероксиде. Липидни хидропероксиди су веома нестабилни и лако се разлажу до секундарних производа, као што су алдехиди (као што је 4-хидрокси-2,3-ноненал) и малондиалдехиди (МДА). Изопростани су још једна група производа пероксидације липида који се стварају пероксидацијом арахидонске киселине и за које је такође утврђено да су повишени у плазми и кондензатима даха код астматичара.34,35 Пероксидација липида нарушава интегритет ћелијских мембрана и доводи до преуређења структура мембране.
Водоник пероксид, супероксидни радикал, оксидовани глутатион (ГССГ), МДА, изопростани, карбонили и нитротирозин могу се лако мерити из плазме, крви или узорака бронхоалвеоларног испирања као биомаркери оксидације стандардизованим тестовима.
Егзогени извор оксиданата
Дим цигарете
Дим цигарета садржи многе оксидансе и слободне радикале и органска једињења, као што су супероксид и азот оксид.36 Поред тога, удисање дима цигарета у плућа такође активира неке ендогене механизме, као што је накупљање неутрофила и макрофага, који додатно повећавају оксидациону повреду. .
Изложеност озону
Изложеност озону може изазвати пероксидацију липида и индуковати прилив неутрофила у епител дисајних путева. Краткотрајно излагање озону такође изазива ослобађање медијатора упале, као што су МПО, еозинофилни катјонски протеини, као и лактат дехидрогеназа и албумин.37 Чак и код здравих субјеката, изложеност озону изазива смањење плућних функција.38 Цхо и сарадници39 су показали да честице (мешавина чврстих честица и течних капљица суспендованих у ваздуху) катализују редукцију кисеоника.
Хиперокиа
Хипероксија се односи на стање већег нивоа кисеоника од нормалног парцијалног притиска кисеоника у плућима или другим телесним ткивима. То доводи до веће производње реактивних врста кисеоника и азота.40,41
Јонизујућег зрачења
Јонизујуће зрачење, у присуству О2, претвара хидроксилни радикал, супероксид и органске радикале у водоник пероксид и органске хидропероксиде. Ове хидропероксидне врсте реагују са редокс активним јонима метала, као што су Фе и Цу, преко Фентонове реакције и на тај начин индукују оксидативни стрес.42,43 Нараианан ет ал44 су показали да фибробласти који су били изложени алфа честицама имају значајно повећање интрацелуларног О2 и Х2О2. производња преко НАДПХ оксидазе везане за плазма мембрану.2 Молекули за трансдукцију сигнала, као што су киназа 44 и 1 регулисана екстрацелуларним сигналом (ЕРК2/1), ц-Јун Н-терминална киназа (ЈНК) и п2, и фактори транскрипције, као нпр. активатор протеин-38 (АП-1), нуклеарни фактор-кБ (НФ-кБ) и п1 се активирају, што резултира експресијом гена повезаних са одговором на зрачење.53�45 Ултраљубичасти А (УВА) фотони покрећу оксидативне реакције ексцитацијом ендогених фотосензибилизатора, као што су порфирини, НАДПХ оксидаза и рибофлавини. 50-Око-8-дихидрогуанин (7,8-окоГуа) је главни УВА-посредовани производ оксидације ДНК који настаје оксидацијом ОХ радикала, 8-електронских оксиданата и синглетног кисеоника који углавном реагује са гванином.1 Формирање гванина Показало се да се катјон радикала у изолованој ДНК ефикасно јавља кроз директан ефекат јонизујућег зрачења.51 Након излагања јонизујућем зрачењу, интрацелуларни ниво глутатиона (ГСХ) опада на кратко, али затим поново расте.52,53
Јони тешких метала
Јони тешких метала, као што су гвожђе, бакар, кадмијум, жива, никл, олово и арсен, могу да изазову стварање реактивних радикала и да изазову ћелијско оштећење кроз смањење активности ензима кроз пероксидацију липида и реакцију са нуклеарним протеинима и ДНК.55
Један од најважнијих механизама стварања слободних радикала посредованих металом је реакција Фентонова типа. Супероксидни јон и водоник пероксид могу да ступе у интеракцију са прелазним металима, као што су гвожђе и бакар, преко Хабер-Вајс/Фентонове реакције катализоване металом да би се формирали ОХ радикали.
Поред механизама типа Фентон и Хабер Веисс, одређени јони метала могу директно да реагују са ћелијским молекулима да генеришу слободне радикале, као што су тиол радикали, или индукују ћелијске сигналне путеве. Ови радикали такође могу да реагују са другим молекулима тиола да би створили О22.. О22. се претвара у Х2О2, што узрокује додатно стварање кисеоникових радикала. Неки метали, као што је арсенит, индукују формирање РОС индиректно активацијом система за производњу радикала у ћелијама.56
Арсен је веома токсичан елемент који производи различите РОС, укључујући супероксид (О2 2), синглет кисеоник (1О2), пероксил радикал (РОО), азот оксид (НО), водоник пероксид (Х2О2) и диметиларсинске перокси радикале [( ЦХ3)2АсОО].57�59 Једињења арсена (ИИИ) могу да инхибирају антиоксидативне ензиме, посебно ензиме зависне од ГСХ, као што су глутатион-С-трансферазе (ГСТ), глутатион пероксидаза (ГСХ-Пк) и ГСХ редуктаза, путем везивања - инг на њихове сулфхидрилне (�СХ) групе.60,61
Олово повећава пероксидацију липида.62 Пријављено је значајно смањење активности СОД ткива и ГПк мозга након излагања олову.63,64 Замена цинка, који служи као кофактор за многе ензиме, оловом, доводи до инактивације таквих ензима. Изложеност олову може изазвати инхибицију ГСТ-а утицајем на тиоле ткива.
РОС генерисан реакцијама катализованим металом може да модификује базе ДНК. Три базне супституције, Г / Ц, Г / Т и Ц / Т, могу настати као резултат оксидативног оштећења јонима метала, као што су Фе21, Цу21 и Ни21. Реид и сарадници65 су показали да је Г / Ц претежно произведен од Фе21, док је Ц / Т супституција била са Цу21 и Ни21.
АНТИОКСИДАНТИ
Људско тело је опремљено разним антиоксидансима који служе за противтежу дејству оксиданата. За све практичне сврхе, они се могу поделити у 2 категорије: ензимске (Табела 2) и неензимске (Табела 3).
Ензимски антиоксиданси
Главни ензимски антиоксиданси плућа су СОД (ЕЦ 1.15.1.11), каталаза (ЕЦ 1.11.1.6) и ГСХ-Пк (ЕЦ 1.11.1.9). Поред ових главних ензима, други антиоксиданси, укључујући хем оксигеназу-1 (ЕЦ 1.14.99.3) и редокс протеине, као што су тиоредоксини (ТРКСс, ЕЦ 1.8.4.10), пероксиредоксини (ПРКС, ЕЦ 1.11.1.15) и глутаредоксини , такође је утврђено да играју кључну улогу у плућној антиоксидативној одбрани.
Пошто је супероксид примарни РОС произведен из различитих извора, његова дисмутација помоћу СОД је од примарне важности за сваку ћелију. Сва 3 облика СОД, односно ЦуЗн-СОД, Мн-СОД и ЕЦ-СОД, широко су изражена у људским плућима. Мн-СОД је локализован у матриксу митохондрија. ЕЦ-СОД је првенствено локализован у екстрацелуларном матриксу, посебно у областима које садрже велике количине колагених влакана типа И и око плућних и системских судова. Такође је откривен у бронхијалном епителу, алвеоларном епителу и алвеоларним макрофагима.66,67 Све у свему, сматра се да ЦуЗн-СОД и Мн-СОД делују као велики сакупљачи супероксидних радикала. Релативно висок ниво ЕЦ-СОД у плућима са његовим специфичним везивањем за компоненте екстрацелуларног матрикса може представљати основну компоненту заштите плућног матрикса.68
Х2О2 који се производи деловањем СОД или деловањем оксидаза, као што је ксантин оксидаза, редукује се у воду помоћу каталазе и ГСХ-Пк. Каталаза постоји као тетрамер састављен од 4 идентична мономера, од којих сваки садржи хем групу на активном месту. Разградња Х2О2 се постиже конверзијом између 2 конформације каталазе-ферикаталазе (гвожђе координисано са водом) и једињења И (гвожђе у комплексу са атомом кисеоника). Каталаза такође везује НАДПХ као редукциони еквивалент да би спречила оксидативну инактивацију ензима (формирање једињења ИИ) Х2О2 док се редукује у воду.69
Ензими у редокс циклусу одговорни за редукцију Х2О2 и липидних хидропероксида (генерисаних као резултат пероксидације мембранских липида) укључују ГСХ-Пкс.70 ГСХ-Пкс су породица тетрамерних ензима који садрже јединствену аминокиселину селеноцистеин унутар активна места и користе тиоле ниске молекуларне тежине, као што је ГСХ, да редукују Х2О2 и липидне пероксиде у њихове одговарајуће алкохоле. Описана су четири ГСХ-Пкс кодирана различитим генима: ГСХ-Пк-1 (ћелијски ГСХ-Пк) је свеприсутан и смањује Х2О2 и пероксиде масних киселина, али не и естерификоване пероксил липиде.71 Естерификовани липиди се редукују ГСХ везаним за мембрану. -Пк-4 (фосфолипидни хидропероксид ГСХ-Пк), који може да користи неколико различитих тиола ниске молекулске тежине као редукционих еквивалената. ГСХ-Пк-2 (гастроинтестинални ГСХ-Пк) је локализован у гастроинтестиналним епителним ћелијама где служи за смањење пероксида у исхрани.72 ГСХ-Пк-3 (екстрацелуларни ГСХ-Пк) је једини члан породице ГСХ-Пк који живи у екстрацелуларни одељак и верује се да је један од најважнијих екстрацелуларних антиоксидативних ензима код сисара. Од њих, екстрацелуларни ГСХ-Пк се највише истражује у људским плућима.73
Поред тога, одлагање Х2О2 је блиско повезано са неколико ензима који садрже тиол, наиме, ТРКСс (ТРКС1 и ТРКС2), тиоредоксин редуктазе (ЕЦ 1.8.1.9) (ТРРс), ПРКСс (који су тиоредоксин пероксидазе) и глута74.
Два ТРКС и ТРР су окарактерисана у људским ћелијама, постоје и у цитосолу и у митохондријама. У плућима, ТРКС и ТРР се експримирају у бронхијалном и алвеоларном епителу и макрофагима. У људским ћелијама је пронађено шест различитих ПРКС-ова, који се разликују по својој ултраструктурној компартментализацији. Експерименталне студије су откриле значај ПРКС ВИ у заштити алвеоларног епитела. Људска плућа експримирају све ПРКС у бронхијалном епителу, алвеоларном епителу и макрофагима.75 Недавно је откривено да ПРКС В функционише као пероксинитрит редуктаза,76 што значи да може функционисати као потенцијално заштитно једињење у развоју РОС-посредоване повреде плућа. .77
Заједничко за ове антиоксиданте је потреба за НАДПХ као редукционим еквивалентом. НАДПХ одржава каталазу у активном облику и користи се као кофактор од стране ТРКС и ГСХ редуктазе (ЕЦ 1.6.4.2), која претвара ГССГ у ГСХ, ко-супстрат за ГСХ-Пкс. Интраћелијски НАДПХ, заузврат, настаје редукцијом НАДП1 помоћу глукоза-6-фосфат дехидрогеназе, првог ензима који ограничава брзину пентоза фосфатног пута, током конверзије глукоза-6-фосфата у 6-фосфоглуконолактон. Генерисањем НАДПХ, глукоза-6-фосфат дехидрогеназа је критична детерминанта пуферског капацитета цитосолног ГСХ (ГСХ/ГССГ) и стога се може сматрати есенцијалним, регулаторним антиоксидативним ензимом.78,79
ГСТ (ЕЦ 2.5.1.18), друга породица антиоксидативних ензима, инактивирају секундарне метаболите, као што су незасићени алдехиди, епоксиди и хидропероксиди. Описане су три главне породице ГСТ: цитосолни ГСТ, митохондријски ГСТ,80,81 и микрозомални ГСТ повезан са мембраном који има улогу у метаболизму еикосаноида и ГСХ.82 Седам класа цитосолног ГСТ је идентификовано код сисара, означених као Алпха, Му, Пи, Сигма, Тхета, Омега и Зета.83�86 Током услова без стреса, ГСТ класе Му и Пи интерагују са киназама Аск1 и ЈНК, респективно, и инхибирају ове киназе.87�89 Показало се да се ГСТП1 дисоцира од ЈНК као одговор на оксидативни стрес.89 ГСТП1 такође физички реагује са ПРКС ВИ и доводи до опоравка активности ензима ПРКС путем глутатионилације оксидованог протеина.90
Неензимски антиоксиданси
Неензимски антиоксиданси укључују једињења мале молекулске тежине, као што су витамини (витамини Ц и Е), б-каротен, мокраћна киселина и ГСХ, трипептид (Лг-глутамил-Л-цистеинил-Л-глицин) који садржи тиол ( сулфхидрил) група.
Витамин Ц (аскорбинска киселина)
Витамин Ц растворљив у води (аскорбинска киселина) обезбеђује интрацелуларни и екстрацелуларни антиоксидативни капацитет водене фазе првенствено уклањањем слободних радикала кисеоника. Конвертује слободне радикале витамина Е назад у витамин Е. Показало се да се његови нивои у плазми смањују са годинама.91,92
витамин Е (а-токоферол)
Витамин Е растворљив у липидима концентрисан је у хидрофобном унутрашњем месту ћелијске мембране и представља главну одбрану од повреде мембране изазване оксидантима. Витамин Е донира електрон пероксил радикалу, који се производи током пероксидације липида. а-токоферол је најактивнији облик витамина Е и главни антиоксиданс везан за мембрану у ћелији. Витамин Е покреће апоптозу ћелија рака и инхибира формације слободних радикала.93
Глутатион
ГСХ је веома богат у свим ћелијским деловима и главни је растворљиви антиоксидант. Однос ГСХ/ГССГ је главна детерминанта оксидативног стреса. ГСХ показује своје антиоксидативне ефекте на неколико начина.94 Детоксикује водоник пероксид и липидне пероксиде деловањем ГСХ-Пк. ГСХ донира свој електрон Х2О2 да би га редуковао у Х2О и О2. ГССГ се поново редукује у ГСХ помоћу ГСХ редуктазе која користи НАД(П)Х као донора електрона. ГСХ-Пкс су такође важни за заштиту ћелијске мембране од пероксидације липида. Редуковани глутатион донира протоне мембранским липидима и штити их од напада оксиданса.95
ГСХ је кофактор за неколико ензима за детоксикацију, као што су ГСХ-Пк и трансфераза. Има улогу у претварању витамина Ц и Е у њихове активне облике. ГСХ штити ћелије од апоптозе интеракцијом са проапоптотичким и антиапоптотичким сигналним путевима.94 Такође регулише и активира неколико фактора транскрипције, као што су АП-1, НФ-кБ и Сп-1.
каротеноиди (б-каротен)
Каротеноиди су пигменти који се налазе у биљкама. Примарно, утврђено је да б-каротен реагује са пероксил (РОО), хидроксил (ОХ) и супероксид (О22.) радикалима.96 Каротеноиди показују своје антиоксидативне ефекте при ниском парцијалном притиску кисеоника, али могу имати прооксидантне ефекте при већем кисеонику. концентрације.97 И каротеноиди и ретиноична киселина (РА) су у стању да регулишу факторе транскрипције.98 б-каротен инхибира активацију НФ-кБ изазвану оксидантима и производњу интерлеукина (ИЛ)-6 и туморске некрозе-а. Каротеноиди такође утичу на апоптозу ћелија. Антипролиферативни ефекти РА су приказани у неколико студија. Овај ефекат РА је посредован углавном рецепторима ретиноичне киселине и варира међу типовима ћелија. У ћелијама карцинома дојке, показало се да рецептор ретиноичне киселине изазива инхибицију раста изазивањем заустављања ћелијског циклуса, апоптозе или обоје.99,100
ЕФЕКАТ ОКСИДАТИВНОГ СТРЕСА: ГЕНЕТСКИ, ФИЗИОЛОШКИ И БИОХЕМИЈСКИ МЕХАНИЗМИ
Оксидативни стрес се јавља када је равнотежа између антиоксиданата и РОС поремећена због исцрпљивања антиоксиданата или акумулације РОС. Када дође до оксидативног стреса, ћелије покушавају да се супротставе оксидативним ефектима и поврате редокс равнотежу активацијом или утишавањем гена који кодирају одбрамбене ензиме, факторе транскрипције и структурне протеине.101,102 Однос између оксидованог и редукованог глутатиона (2ГСХ/ГССГ) је један. важних детерминанти оксидативног стреса у телу. Већа производња РОС у телу може променити структуру ДНК, резултирати модификацијом протеина и липида, активацијом неколико фактора транскрипције изазваних стресом и производњом проинфламаторних и антиинфламаторних цитокина.
Ефекти оксидативног стреса на ДНК
РОС може довести до модификација ДНК на неколико начина, што укључује деградацију база, једно- или дволанчане ломове ДНК, пуринске, пиримидинске или модификације везане за шећер, мутације, делеције или транслокације и унакрсно повезивање са протеинима. Већина ових ДНК модификација (слика 1) су веома релевантне за канцерогенезу, старење и неуродегенеративне, кардиоваскуларне и аутоимуне болести. Дувански дим, редокс метали и нередокс метали, као што су гвожђе, кадмијум, хром и арсен, такође су укључени у карциногенезу и старење тако што стварају слободне радикале или се везују за тиолне групе. Формирање 8-ОХ-Г је најпознатије оштећење ДНК које настаје оксидативним стресом и потенцијални је биомаркер за карциногенезу.
Промоторски региони гена садрже консензус секвенце за факторе транскрипције. Ова места везивања фактора транскрипције садрже секвенце богате ГЦ које су подложне нападима оксиданса. Формирање 8-ОХ-Г ДНК на местима везивања фактора транскрипције може модификовати везивање транскрипционих фактора и на тај начин променити експресију сродних гена као што је показано за циљне секвенце АП-1 и Сп-1 Поред 103-ОХ-Г, Такође се показало да 8-цикло-8,59-деоксиаденозин (цикло-дА) инхибира транскрипцију са репортерског гена у ћелијском систему ако се налази у ТАТА кутији.29 ТАТА-везујући протеин иницира транскрипцију променом савијања ДНК . Везивање ТАТА-везујућег протеина може бити нарушено присуством цикло-дА.
Оксидативни стрес изазива нестабилност микросателитских (кратких тандем понављања) региона. Редокс активни јони метала, хидроксилни радикали повећавају нестабилност микросателита.105 Иако ћелије могу лако толерисати једноланчане ломове ДНК узроковане повредом оксиданса, дволанчани прекиди ДНК изазвани јонизујућим зрачењем могу бити значајна претња за опстанак ћелије.106
Метилација на ЦпГ острвима у ДНК је важан епигенетски механизам који може довести до утишавања гена. Оксидација 5-МеЦит у 5-хидроксиметил урацил (5-ОХМеУра) може се десити преко реакција деаминације/оксидације тимина или интермедијера 5-хидроксиметил цитозина.107 Поред модулације експресије гена, чини се да метилација ДНК такође утиче на организацију хроматина.108 Аберантни обрасци метилације ДНК изазвани оксидативним нападима такође утичу на активност поправке ДНК.
Ефекти оксидативног стреса на липиде
РОС може индуковати пероксидацију липида и пореметити двослој мембране липида који може инактивирати рецепторе везане за мембрану и ензиме и повећати пермеабилност ткива.109 Производи пероксидације липида, као што су МДА и незасићени алдехиди, способни су да инактивирају многе ћелијске протеине формирањем укрштања протеина. -везе.110�112 4-Хидроки-2-ноненал изазива исцрпљивање интрацелуларног ГСХ и индукује производњу пероксида,113,114 активира рецептор епидермалног фактора раста,115 и индукује производњу фибронектина.116 Производи пероксидације липида, као што су супстанци изобарбитне киселине и реактивне киселине , коришћени су као индиректни биомаркери оксидативног стреса, а повећани нивои су приказани у издисаном кондензату или бронхоалвеоларној течности за испирање или плућима пацијената са хроничном опструктивном болешћу плућа или пушача.117�119
Ефекти оксидативног стреса на протеине
РОС може изазвати фрагментацију пептидног ланца, промену електричног набоја протеина, умрежавање протеина и оксидацију специфичних амино киселина и стога довести до повећане осетљивости на протеолизу разградњом специфичних протеаза.120 Остаци цистеина и метионина у протеинима су посебно подложнији оксидацији.121 Оксидација сулфхидрилних група или остатака метионина протеина изазива конформационе промене, одвијање протеина и деградацију.8,121�123 Ензими који имају метале на или близу својих активних места су посебно осетљивији на оксидацију катализовану металом. Показало се да оксидативна модификација ензима инхибира њихову активност.124,125
У неким случајевима може доћи до специфичне оксидације протеина. На пример, метионин се може оксидовати метионин сулфоксидом126 и фенилаланином у о-тирозин127; сулфхидрилне групе се могу оксидовати да би се формирале дисулфидне везе;128 и карбонил групе могу бити уведене у бочне ланце протеина. Гама зраци, оксидација катализована металом, ХОЦл и озон могу изазвати стварање карбонилних група.129
Ефекти оксидативног стреса на трансдукцију сигнала
РОС може индуковати експресију неколико гена укључених у трансдукцију сигнала.1,130 Висок однос за ГСХ/ГССГ је важан за заштиту ћелије од оксидативног оштећења. Поремећај овог односа изазива активацију редокс осетљивих транскрипционих фактора, као што су НФ-кБ, АП-1, нуклеарни фактор активираних Т ћелија и хипоксијом индуцибилни фактор 1, који су укључени у инфламаторни одговор. Активација транскрипционих фактора преко РОС-а се постиже каскадама трансдукције сигнала које преносе информације споља у унутрашњост ћелије. Рецептори тирозин киназе, већина рецептора фактора раста, као што су рецептор епидермалног фактора раста, рецептор васкуларног ендотелног фактора раста и рецептор за фактор раста који потиче од тромбоцита, протеинске тирозин фосфатазе и серин/треонин киназе су мете РОС.131�133 Екстраћелијске сигнално регулисане киназе, ЈНК и п38, које су чланови породице протеин киназа активираних митогеном и укључене у неколико процеса у ћелији укључујући пролиферацију, диференцијацију и апоптозу, такође могу бити регулисане оксидантима.
У условима оксидативног стреса, цистеински остаци на месту везивања за ДНК ц-Јун, неке АП-1 подјединице и инхибиторна кБ киназа подлежу реверзибилној С-глутатиолацији. Пријављено је да глутаредоксин и ТРКС играју важну улогу у регулацији сигналних путева осетљивих на редокс, као што су НФ-кБ и АП-1, протеин киназа активирана п38 митогеном и ЈНК.134�137
НФ-кБ се може активирати као одговор на услове оксидативног стреса, као што су РОС, слободни радикали и УВ зрачење.138 Фосфорилација ИкБ ослобађа НФ-кБ и омогућава му да уђе у језгро како би активирао транскрипцију гена.139 Бројни киназе имају пријављено је да фосфорилише ИкБ на серинским остацима. Ове киназе су мете оксидативних сигнала за активацију НФ-кБ.140 Редукциони агенси појачавају везивање НФ-кБ ДНК, док оксидациони агенси инхибирају ДНК везивање НФ-кБ. ТРКС може имати 2 супротна дејства у регулацији НФ-кБ: у цитоплазми, блокира деградацију ИкБ и инхибира активацију НФ-кБ, али појачава везивање НФ-кБ ДНК у језгру.141 Активација НФ-кБ деградацијом повезане са оксидацијом ИкБ доводи до активације неколико гена повезаних са антиоксидативном одбраном. НФ-кБ регулише експресију неколико гена који учествују у имунолошком одговору, као што су ИЛ-1б, ИЛ-6, фактор некрозе тумора-а, ИЛ-8 и неколико молекула адхезије.142,143 НФ-кБ такође регулише ангиогенезу и пролиферацију и диференцијација ћелија.
АП-1 је такође регулисан редокс стањем. У присуству Х2О2, неки метални јони могу изазвати активацију АП-1. Повећање односа ГСХ/ГССГ појачава везивање АП-1, док ГССГ инхибира ДНК везивање АП-1.144 ДНК везивање Фос/Јун хетеродимера се повећава смањењем једног очуваног цистеина у домену за везивање ДНК сваког од протеини,145 док ДНК везивање АП-1 може бити инхибирано ГССГ-ом у многим типовима ћелија, што сугерише да формирање дисулфидне везе помоћу цистеинских остатака инхибира везивање АП-1 ДНК.146,147 Трансдукција сигнала путем оксидативног стреса је сажета на слици 2.
ЗАКЉУЧАК
Оксидативни стрес може настати услед прекомерне производње РОС метаболичким реакцијама које користе кисеоник и померају равнотежу између оксиданс/антиоксиданс статусе у корист оксиданата. РОС се производе метаболичким активностима ћелија и факторима животне средине, као што су загађивачи ваздуха или дим цигарета. РОС су високо реактивни молекули због неспарених електрона у својој структури и реагују са неколико биолошких макромолекула у ћелији, као што су угљени хидрати, нуклеинске киселине, липиди и протеини, и мењају њихове функције. РОС такође утиче на експресију неколико гена повећањем регулације транскрипционих фактора осетљивих на редокс и ремоделирањем хроматина преко промене ацетилације/деацетилације хистона. Регулација редокс стања је критична за виталност ћелије, активацију, пролиферацију и функцију органа.
РЕФЕРЕНЦЕ
1. Валко М, Рходес ЦЈ, Монцол Ј, Изаковић М, Мазур М. Слободни радикали, метали и антиоксиданси у раку изазваном оксидативним стресом. Цхем Биол Интерацт. 2006;160:1�40.
2. Халливелл Б, Гуттериџ ЈМЦ. Слободни радикали у биологији и медицини. 3рд ед. Њујорк: Окфорд Университи Пресс; 1999.
3. Марнетт Љ. Пероксидација липида и оштећење ДНК малондиалдехидом. Мутат Рес. 1999;424:83�95.
4. Сиемс ВГ, Груне Т, Естербауер Х. Формирање 4-Хидрокиноненала током исхемије и реперфузије танког црева пацова. � Лифе Сци. 1995;57:785�789.
5. Стадтман ЕР. Улога оксидативних врста у старењу. Цурр Мед Цхем. 2004;11:1105�1112.
6. Ванг МИ, Дхингра К, Хиттелман ВН, Лиехр ЈГ, деАндраде М, Ли ДХ. Претпостављени адукти ДНК малондиалдехида изазвани пероксидацијом липида у ткивима људске дојке. Цанцер Епидемиол Биомаркерс Прев. 1996;5:705�710.
7. Јеннер П. Оксидативни стрес код Паркинсонове болести. Анн Неурол. 2003;53: С26�С36.
8. Лирас Л, Цаирнс Њ, Јеннер А, Јеннер П, Халливелл Б. Процена оксидативног оштећења протеина, липида и ДНК у мозгу пацијената са Алцхајмеровом болешћу. Ј Неуроцхем. 1997;68:2061�2069.
9. Саире ЛМ, Смитх МА, Перри Г. Хемија и биохемија оксидативног стреса у неуродегенеративним болестима. Цурр Мед Цхем. 2001;8:721�738.
10. Тосхнивал ПК, Зарлинг ЕЈ. Докази за повећану пероксидацију липида код мултипле склерозе. Неуроцхем Рес. 1992;17:205�207.
11. Дхалла НС, Темсах РМ, Неттицадан Т. Улога оксидативног стреса у кардиоваскуларним болестима. Ј Хипертенс. 2000;18:655�673.
12. Каспарова С, Брезова В, Валко М, Хорецки Ј, Млинарик В, ет ал. Проучавање оксидативног стреса у моделу хроничне хипоперфузије мозга код пацова. Неуроцхем Инт. 2005;46:601�611.
13. Керр С, Броснан МЈ, МцИнтире М, Реид ЈЛ, Доминицзак АФ, Хамилтон ЦА. Производња супероксид ањона је повећана у моделу генетске хипертензије: улога ендотела. Хипертензија. 1999;33:1353�1358.
14. Кукреја РЦ, Хесс МЛ. Систем слободних радикала кисеоника: од једначина преко интеракција мембранских протеина до кардиоваскуларних повреда и заштите. Цардиовасц Рес. 1992;26:641�655.
15. Асами С, Манабе Х, Мииаке Ј, Тсурудоме И, Хирано Т, ет ал. Пушење цигарета изазива повећање оксидативног оштећења ДНК, 8-хидроксидеоксигуанозина, у централном месту људских плућа. Карциногенеза. 1997;18:1763�1766.
16. Андреадис АА, Хазен СЛ, Цомхаир СА, Ерзурум СЦ. Оксидативни и нитрозативни догађаји код астме. Фрее Радиц Биол Мед. 2003;35:213�225.
17. Цомхаир СА, Рицци КС, Арролига М, Лара АР, Двеик РА, ет ал. Корелација системског недостатка супероксид дисмутазе са опструкцијом протока ваздуха код астме. Ам Ј Респир Црит Царе Мед. 2005;172:306�313.
18. Цомхаир СА, Ксу В, Гхосх С, Тхунниссен ФБ, Алмасан А, ет ал. Инактивација супероксид дисмутазе у патофизиологији ремоделирања и реактивности дисајних путева астме. Ам Ј Патхол. 2005;166:663�674.
19. Дут Р, Диздар ЕА, Бирбен Е, Сацкесен Ц, Соиер ОУ, Беслер Т, Калаици О. Оксидативни стрес и његове детерминанте у дисајним путевима деце са астмом. алергија. 2008;63:1605�1609.
20. Ерцан Х, Бирбен Е, Диздар ЕА, Кескин О, Карааслан Ц, ет ал. Оксидативни стрес и генетске и епидемиолошке детерминанте оксидативне повреде у дечјој астми. Ј Аллерги Цлин Иммунол. 2006;118:1097�1104.
21. Фитзпатрицк АМ, Теагуе ВГ, Холгуин Ф, Иех М, Бровн ЛА. Програм истраживања тешке астме. Хомеостаза глутатиона у дисајним путевима је промењена код деце са тешком астмом: доказ оксидативног стреса. Ј Аллерги Цлин Иммунол. 2009;123:146�152.
22. Миллер ДМ, Буеттнер ГР, Ауст СД. Прелазни метали као катализатори реакција „аутооксидације“. Фрее Радиц Биол Мед. 1990;8:95�108.
23. Дупуи Ц, Вирион А, Охаион Р, Каниевски Ј, Дме Д, Поммиер Ј. Механизам формирања водоник пероксида катализованог НАДПХ оксидазом у тироидној плазма мембрани. Ј Биол Цхем. 1991;266:3739�3743.
24. Грангер ДН. Улога ксантин оксидазе и гранулоцита у исхемијској перфузионој повреди. Ам Ј Пхисиол. 1988;255:Х1269�Х1275.
25. Фентон ХЈХ. Оксидација винске киселине у присуству гвожђа. Ј Цхем Соц. 1984;65:899�910.
26. Хабер Ф, Веисс ЈЈ. Каталитичко разлагање водоник пероксида солима гвожђа. Проц Р Соц Лонд Сер А. 1934;147:332�351.
27. Лиоцхев СИ, Фридовицх И. Хабер-Веисс циклус 70 година касније: алтернативни поглед. Редок Реп. 2002;7:55�57.
28. Клебанофф СЈ. Мијелопероксидаза: пријатељ и непријатељ. Ј Леукоц Биол. 2005;77:598�625.
29. Вхитеман М, Јеннер А, Халливелл Б. Модификације базе изазване хипохлорном киселином у изолованој ДНК тимуса телета. Цхем Рес Токицол. 1997;10:1240�1246.
30. Кулцхарик ПА, Хеинецке ЈВ. Хипохлорна киселина коју производи мијелопероксидазни систем људских фагоцита индукује ковалентне унакрсне везе између ДНК и протеина. Биохемија. 2001;40:3648�3656.
31. Бреннан МЛ, Ву В, Фу Кс, Схен З, Сонг В, ет ал. Прича о две контроверзе: дефинисању улоге пероксидаза у формирању нитротирозина ин виво помоћу мишева са недостатком еозинофил пероксидазе и мијелопероксидазе, и природе реактивних азотних врста које стварају пероксидаза. Ј Биол Цхем. 2002;277:17415�17427.
32. Дензлер КЛ, Борцхерс МТ, Цросби ЈР, Циеслевицз Г, Хинес ЕМ, ет ал. Екстензивна дегранулација еозинофила и оксидација протеина дисајних путева посредована пероксидазом се не јављају у моделу плућне упале код миша који изазива овалбумин. Ј Иммунол. 2001;167:1672�1682.
33. ван Дален ЦЈ, Винтербоурн ЦЦ, Сентхилмохан Р, Кеттле АЈ. Нитрит као супстрат и инхибитор мијелопероксидазе. Импликације за нитрацију и производњу хипохлорне киселине на местима упале. Ј Биол Цхем. 2000;275:11638�11644.
34. Воод ЛГ, Фитзгералд ДА, Гибсон ПГ, Цоопер ДМ, Гарг МЛ. Пероксидација липида, одређена изопростанима у плазми, повезана је са тежином болести код благе астме. Липиди. 2000;35:967�974.
35. Монтусцхи П, Цорради М, Циабаттони Г, Нигхтингале Ј, Кхаритонов СА, Барнес ПЈ. Повећан 8-изопростан, маркер оксидативног стреса, у издахнутом кондензату пацијената са астмом. Ам Ј Респир Црит Царе Мед. 1999;160:216�220.
36. Цхурцх ДФ, Приор ВА. Слободно-радикална хемија дима цигарета и њене токсиколошке импликације. Перспектива здравља животне средине. 1985;64:111�126.
37. Хилтерманн ЈТ, Лапперре ТС, ван Брее Л, Стееренберг ПА, Брахим ЈЈ, ет ал. Упала изазвана озоном процењена у спутуму и течности за испирање бронха код астматичара: ново неинвазивно средство у епидемиолошким студијама о загађењу ваздуха и астми. Фрее Радиц Биол Мед. 1999;27:1448�1454.
38. Нигхтингале ЈА, Рогерс ДФ, Барнес ПЈ. Утицај озона који се удише на издахнути азотни оксид, функцију плућа и индуковани спутум код нормалних и астматичара. Торакс. 1999;54:1061�1069.
39. Цхо АК, Сиоутас Ц, Мигуел АХ, Кумагаи И, Сцхмитз ДА, ет ал. Редок активност честица у ваздуху на различитим локацијама у басену Лос Анђелеса. Енвирон Рес. 2005;99:40�47.
40. Цомхаир СА, Тхомассен МЈ, Ерзурум СЦ. Диференцијална индукција екстрацелуларне глутатион пероксидазе и азот оксид синтазе 2 у дисајним путевима здравих особа изложених 100% О(2) или диму цигарета. Ам Ј Респир Целл Мол Биол. 2000;23:350�354.
41. Маттхаи МА, Геисер Т, Маталон С, Исцхиропоулос Х. Оксидант-посредована повреда плућа у акутном респираторном дистрес синдрому. Црит Царе Мед. 1999;27:2028�2030.
42. Биаглов ЈЕ, Митцхелл ЈБ, Хелд К. Значај пероксида и супероксида у рендгенском одговору. Инт Ј Радиат Онцол Биол Пхис. 1992;22:665�669.
43. Цхиу СМ, Ксуе ЛИ, Фриедман ЛР, Олеиницк НЛ. Сензибилизација места везивања нуклеарног матрикса на јонизујуће зрачење посредована јонима бакра. Биохемија. 1993;32:6214�6219.
44. Нараианан ПК, Гоодвин ЕХ, Лехнерт БЕ. Алфа честице иницирају биолошку производњу супероксидних ањона и водоник пероксида у људским ћелијама. Цанцер Рес. 1997;57:3963�3971.
45. Туттле СВ, Варнес МЕ, Митцхелл ЈБ, Биаглов ЈЕ. Осетљивост на хемијске оксиданте и зрачење у ЦХО ћелијским линијама са недостатком активности оксидативног пентозног циклуса. Инт Ј Радиат Онцол Биол Пхис. 1992;22: 671�675.
46. Гуо Г, Иан-Сандерс И, Лин-Цоок БД, Ванг Т, Тамае Д, ет ал. манган
експресија гена посредована супероксид дисмутазом у зрачењу
адаптивни одговори. Мол Целл Биол. 2003;23:2362�2378.
47. Аззам ЕИ, де Толедо СМ, Спитз ДР, Литтле ЈБ. Оксидативни метаболизам
модулира трансдукцију сигнала и формирање микронуклеуса код посматрача
ћелије из а-честица озрачених нормалних хуманих фибробласта. Цанцер Рес.
2002;62:5436�5442.
48. Леацх ЈК, Ван Туиле Г, Лин ПС, Сцхмидт-Уллрицх Р, Миккелсен РБ.
Генерација реактивних изазвана јонизујућим зрачењем, зависна од митохондрија
кисеоник/азот. Цанцер Рес. 2001;61:3894�3901.
49. Дент П, Иацоуб А, Фисхер ПБ, Хаган МП, Грант С. МАПК путеви у
реакције на зрачење. Онкоген. 2003;22:5885�5896.
50. Веи СЈ, Ботеро А, Хирота К, Брадбури ЦМ, Марковина С, ет ал. Тиоредоксин
нуклеарна транслокација и интеракција са редокс фактором-1 активира фактор транскрипције АП-1 као одговор на јонизујуће зрачење. Цанцер Рес. 2000;60:6688�6695.
51. Кадет Ј, Доуки Т, Гаспарутто Д, Раванат ЈЛ. Оксидативна оштећења ДНК: формирање, мерење и биохемијске карактеристике. Мутат Рес. 2003;531:5�23.
52. Иокоиа А, Цунниффе СМ, О'Неилл П. Ефекат хидратације на индукцију прекида ланаца и лезија база у филмовима плазмидне ДНК гамарадијацијом. Ј Ам Цхем Соц. 2002;124:8859�8866.
53. Јанссен ИМ, Ван Хоутен Б, Борм ПЈ, Моссман БТ. Реакције ћелија и ткива на оксидативна оштећења. Лаб Инвест. 1993;69:261�274.
54. Иванага М, Мори К, Иида Т, Урата И, Матсуо Т, ет ал. Индукција гама глутамилцистеин синтетазе зависна од нуклеарног фактора капа Б јонизујућим зрачењем у Т98Г ћелијама хуманог глиобластома. Фрее Радиц Биол Мед. 1998;24:1256�1268.
55. Стохс СЈ, Багцхи Д. Оксидативни механизми у токсичности металних јона. Фрее Радиц Биол Мед. 1995;18:321�336.
56. Леонард СС, Харрис ГК, Схи Кс. Металом индуковани оксидативни стрес и трансдукција сигнала. Фрее Радиц Биол Мед. 2004;37:1921�1942.
57. Схи Х, Схи Кс, Лиу КЈ. Оксидативни механизам токсичности арсена и карциногенезе. Мол Целл Биоцхем. 2004;255:67�78.
58. Пи Ј, Хоригуцхи С, Сун И, Никаидо М, Схимојо Н, Хаиасхи Т. Потенцијални механизам за оштећење формирања азотног оксида узрокованог продуженим оралним излагањем арсенату код зечева. Фрее Радиц Биол Мед.2003;35:102�113.
59. Рин К, Кавагуцхи К, Иаманака К, Тезука М, Оку Н, Окада С. Прекиди ДНК нити изазвани диметиларсинском киселином, метаболитом неорганског арсеника, снажно су појачани супероксидним ањонским радикалима. Биол Пхарм Булл. 1995;18:45�58.
60. Ваалкес МП, Лиу Ј, Вард ЈМ, Диван ЛА. Механизми који леже у основи карциногенезе арсена: преосетљивост мишева изложених неорганском арсену током гестације. Токицологи. 2004;198:31�38.
61. Сцхиллер ЦМ, Фовлер БА, Воодс ЈС. Ефекти арсена на активацију пируват дехидрогеназе. Перспектива здравља животне средине. 1977;19:205�207.
62. Монтерио ХП, Бецхара ЕЈХ, Абдалла ДСП. Учешће слободних радикала у неуролошким порфиријама и тровању оловом. Мол Целл Биоцхем. 1991;103:73�83.
63. Трипатхи РМ, Рагхунатх Р, Махапатра С. Олово у крви и његов утицај на нивое Цд, Цу, Зн, Фе и хемоглобина код деце. Сци Тотал Енвирон. 2001;277:161�168.
64. Нехру Б, Дуа Р. Ефекат дијететског селена на неуротоксичност олова. Ј Енвирон Патхол Токицол Онцол. 1997;16:47�50.
65. Реид ТМ, Феиг ДИ, Лоеб ЛА. Мутагенеза радикалима кисеоника изазваним металом. Перспектива здравља животне средине. 1994;102(суппл 3):57.
66. Киннула ВЛ, Црапо ЈД. Супероксид дисмутазе у плућима и плућним болестима људи. Ам Ј Респир Црит Царе Мед. 2003;167:1600�1619.
67. Киннула ВЛ. Производња и разградња метаболита кисеоника током инфламаторних стања у људским плућима. Цурр лек циља на упалну алергију. 2005;4:465�470.
68. Зелко ИН, Мариани ТЈ, Фолз РЈ. Мултигенска породица супероксид дисмутазе: поређење ЦуЗн-СОД (СОД1), Мн-СОД (СОД2) и ЕЦ-СОД (СОД3) генске структуре, еволуција и експресија. Фрее Радиц Биол Мед. 2002;33:337�349.
69. Киркман ХН, Ролфо М, Феррарис АМ, Гаетани ГФ. Механизми заштите каталазе помоћу НАДПХ. Кинетика и стехиометрија. Ј Биол Цхем. 1999;274:13908�13914.
70. Флох Л. Глутатион пероксидаза. Басиц Лифе Сци. 1988;49:663�668.
71. Артхур ЈР. Глутатион пероксидазе. Целл Мол Лифе Сци. 2000;57:1825�1835.
72. Цху ФФ, Доросхов ЈХ, Есвортхи РС. Експресија, карактеризација и дистрибуција ткива нове ћелијске селен-зависне глутатион пероксидазе, ГСХПк-ГИ. Ј Биол Цхем. 1993;268:2571�2576.
73. Цомхаир СА, Бхатхена ПР, Фарвер Ц, Тхунниссен ФБ, Ерзурум СЦ. Екстрацелуларна индукција глутатион пероксидазе у астматичним плућима: доказ редокс регулације експресије у епителним ћелијама људских дисајних путева. ФАСЕБ Ј. 2001;15:70.
74. Громер С, Уриг С, Бецкер К. Тхе тхиоредокин системдфром сциенце то цлиниц. Мед Рес Рев. 2004;24:40�89.
75. Киннула ВЛ, Лехтонен С, Каартеенахо-Виик Р, Лакари Е, П��кк� П, ет ал. Специфична ћелијска експресија пероксиредоксина у хуманој плућној и плућној саркоидози. Торакс. 2002;57:157�164.
76. Дубуиссон М, Вандер Стрицхт Д, Цлиппе А, Етиенне Ф, Наусер Т, ет ал. Хумани пероксиредоксин 5 је пероксинитрит редуктаза. ФЕБС Летт. 2004;571:161�165.
77. Холмгрен А. Антиоксидативна функција тиоредоксинских и глутаредоксинских система. Антиоксид Редок сигнал. 2000;2:811�820.
78. Дицкинсон ДА, Форман ХЈ. Глутатион у одбрани и сигнализацији: лекције из малог тиола. Анн НИ Ацад Сци. 2002;973:488�504.
79. Сиес Х. Глутатион и његова улога у ћелијским функцијама. Фрее Радиц Биол Мед. 1999;27:916�921.
80. Ладнер ЈЕ, Парсонс ЈФ, Рифе ЦЛ, Гиллиланд ГЛ, Армстронг РН. Паралелни еволутивни путеви за глутатион трансферазе: структура и механизам капа ензима митохондријалне класе рГСТК1-1. Биохемија. 2004;43:52�61.
81. Робинсон А, Хуттлеи ГА, Боотх ХС, Боард ПГ. Студије моделирања и биоинформатике људске капа класе глутатион трансферазе предвиђају нову трећу породицу трансфераза са хомологијом са прокариотским 2-хидроксихромен-2-карбоксилат изомеразама. Биоцхем Ј. 2004;379:541-552.
82. Јакобссон ПЈ, Моргенстерн Р, Манцини Ј, Форд-Хутцхинсон А, Перссон Б. Заједничке структурне карактеристике МАПЕГда широко распрострањене суперфамилије протеина повезаних са мембраном са веома дивергентним функцијама у метаболизму еикозаноида и глутатиона. Протеин Сци. 1999;8:689�692.
83. Хаиес ЈД, Пулфорд ДЈ. Фамилија супергена глутатион С-трансферазе: регулација ГСТ-а и допринос изоензима хемопротекцији рака и отпорности на лекове. Црит Рев Биоцхем Мол Биол. 1995;30:445�600.
84. Армстронг РН. Структура, каталитички механизам и еволуција глутатион трансфераза. Цхем Рес Токицол. 1997;10:2�18.
85. Хаиес ЈД, МцЛеллан ЛИ. Глутатион и ензими зависни од глутатиона представљају координирано регулисану одбрану од оксидативног стреса. Фрее Радиц Рес. 1999;31:273�300.
86. Схеехан Д, Меаде Г, Фолеи ВМ, Довд ЦА. Структура, функција и еволуција глутатион трансфераза: импликације за класификацију несисарских чланова древне суперфамилије ензима. Биоцхем Ј. 2001;360:1-16.
87. Цхо СГ, Лее ИХ, Парк ХС, Риоо К, Канг КВ, ет ал. Глутатион С-трансфераза Му модулира сигнале активиране стресом потискивањем киназе која регулише сигнал апоптозе 1. Ј Биол Цхем. 2001;276:12749�12755.
88. Дорион С, Ламберт Х, Ландри Ј. Активација п38 сигналног пута топлотним шоком укључује дисоцијацију глутатион С-трансферазе Му из Аск1. Ј Биол Цхем. 2002;277:30792�30797.
89. Адлер В, Иин З, Фуцхс СИ, Бенезра М, Росарио Л, ет ал. Регулација ЈНК сигнализације ГСТп. ЕМБО Ј. 1999;18:1321-1334.
90. Маневицх И, Феинстеин СИ, Фисхер АБ. Активација антиоксидативног ензима 1-ЦИС пероксиредоксина захтева глутатионилацију посредовану хетеродимеризацијом са пГСТ. Проц Натл Ацад Сци УС А. 2004;101:3780�3785.
91. Бункер ВВ. Слободни радикали, антиоксиданси и старење. Мед Лаб Сци. 1992;49:299�312.
92. Меззетти А, Лапенна Д, Романо Ф, Цостантини Ф, Пиердоменицо СД, ет ал. Системски оксидативни стрес и његов однос са годинама и болешћу. Ј Ам Гериатр Соц. 1996;44:823�827.
93. Вхите Е, Сханнон ЈС, Паттерсон РЕ. Однос између витамина и
употреба суплемената калцијума и рака дебелог црева. Цанцер Епидемиол Биомаркерс Прев. 1997;6:769�774.
94. Маселла Р, Ди Бенедетто Р, Вари Р, Филеси Ц, Гиованнини Ц. Нови механизми природних антиоксидативних једињења у биолошким системима: укљученост глутатиона и ензима повезаних са глутатионом. Ј Нутр Биоцхем. 2005;16:577�586.
95. Цурелло С, Цецони Ц, Биголи Ц, Феррари Р, Албертини А, Гуарниери Ц. Промене у статусу срчаног глутатиона након исхемије и реперфузије. Екпериентиа. 1985;41:42�43.
96. Ел-Агамеи А, Лове ГМ, МцГарвеи ДЈ, Мортенсен А, Пхиллип ДМ, Трусцотт ТГ. Хемија радикала каротеноида и антиоксидативна/прооксидативна својства. Арцх Биоцхем Биопхис. 2004;430:37�48.
97. Рице-Еванс ЦА, Сампсон Ј, Брамлеи ПМ, Холловаи ДЕ. Зашто очекујемо да каротеноиди буду антиоксиданси ин виво? Фрее Радиц Рес. 1997;26:381�398.
98. Нилес РМ. Сигнални путеви у ретиноидној хемопревенцији и лечењу рака. Мутат Рес. 2004;555:81�96.
99. Донато Љ, Нои Н. Суппресија раста карцинома дојке ретиноинском киселином: проапоптотички гени су мете за сигнализацију рецептора ретиноичне киселине и ћелијског протеина ИИ који везује ретиноичну киселину. Цанцер Рес. 2005;65:8193�8199.
100. Ниизума Х, Накамура И, Озаки Т, Наканисхи Х, Охира М, ет ал. Бцл-2 је кључни регулатор за апоптотичку смрт ћелија изазвану ретиноинском киселином у неуробластому. Онкоген. 2006;25:5046�5055.
101. Далтон ТП, Схертзер ХГ, Пуга А. Регулација експресије гена реактивним кисеоником. Анн Рев Пхармацол Токицол. 1999;39:67�101.
102. Сцандалиос ЈГ. Геномски одговори на оксидативни стрес. У: Меиерс РА, ур. Енциклопедија молекуларне биологије ћелије и молекуларне медицине. Вол 5. 2нд ед. Вајнхајм, Немачка: Вилеи-ВЦХ; 2004: 489�512.
103. Гхосх Р, Митцхелл ДЛ. Ефекат оксидативног оштећења ДНК у елементима промотера на везивање фактора транскрипције. Нуклеинске киселине Рес. 1999;27:3213�3218.
104. Мариетта Ц, Гулам Х, Броокс ПЈ. Једна лезија 8-цикло-50-деоксиаденозина у ТАТА кутији спречава везивање ТАТА везујућег протеина и снажно смањује транскрипцију ин виво. Поправка ДНК (Амст). 20;2002:1�967.
105. Јацксон АЛ, Цхен Р, Лоеб ЛА. Индукција нестабилности микросателита
оксидативним оштећењем ДНК. Проц Натл Ацад Сци УС А. 1998;95:12468�12473.
106. Цалдецотт КВ. Интеракције протеин-протеин током поправке једноланчаног прекида ДНК сисара. Биоцхем Соц Транс. 2003;31:247�251.
107. Цооке МС, Еванс МД, Диздароглу М, Лунец Ј. Оксидативно оштећење ДНК: механизми, мутације и болест. ФАСЕБ Ј. 2003;17:1195�1214.
108. Јонес ПЛ, Волффе АП. Односи између организације хроматина и метилације ДНК у одређивању експресије гена. Семин Цанцер Биол. 1999;9:339�347.
109. Гиротти АВ. Механизми пероксидације липида. Ј Фрее Радиц Биол Мед. 1985;1:87�95.
110. Сиу ГМ, Драпер ХХ. Метаболизам малоналдехида ин виво и ин витро. Липиди. 1982;17:349�355.
111. Естербауер Х, Коллер Е, Слее РГ, Костер ЈФ. Могуће учешће продукта пероксидације липида 4-хидроксиноненала у формирању флуоресцентних хромолипида. Биоцхем Ј. 1986;239:405-409.
112. Хагихара М, Нисхигаки И, Масеки М, Иаги К. Промене зависне од старости у нивоима липидног пероксида у фракцијама липопротеина људског серума. Ј Геронтол. 1984;39:269�272.
113. Келлер ЈН, Марк РЈ, Бруце АЈ, Бланц Е, Ротхстеин ЈД, ет ал. 4- Хидроксиноненал, алдехидни производ пероксидације мембранских липида, отежава транспорт глутамата и митохондријалну функцију у синаптозомима. Неуросциенце. 1997;806:85�96.
114. Уцхида К, Схираисхи М, Наито И, Тории И, Накамура И, Осава Т. Активација путева сигнализације стреса крајњим производом пероксидације липида. 4-хидрокси-2-ноненал је потенцијални индуктор производње интрацелуларног пероксида. Ј Биол Цхем. 1999;274:2234�2242.
115. Суц И, Меилхац О, Лајоие-Мазенц И, Вандаеле Ј, Јургенс Г, Салваире Р, Негре-Салваире А. Активација ЕГФ рецептора оксидованим ЛДЛ. ФАСЕБ Ј. 1998;12:665-671.
116. Тсукагосхи Х, Кавата Т, Схимизу И, Исхизука Т, Добасхи К, Мори М. 4-Хидроки-2-ноненал повећава производњу фибронектина ИМР-90 хуманим плућним фибробластима делимично преко активације екстрацелуларног сигнала повезаног са рецептором епидермалног фактора раста- регулисани пут киназе п44/42. Токицол Аппл Пхармацол. 2002;184:127�135.
117. Монтусцхи П, Цоллинс ЈВ, Циабаттони Г, Лаззери Н, Цорради М, Кхаритонов СА, Барнес ПЈ. Издахнути 8-изопростан као ин виво биомаркер оксидативног стреса плућа код пацијената са ХОБП и здравих пушача. Ам Ј Респир Црит Царе Мед. 2000;162:1175�1177.
118. Морисон Д, Рахман И, Ланнан С, МацНее В. Епителна пермеабилност, инфламација и оксидативни стрес у ваздушним просторима пушача. Ам Ј Респир Црит Царе Мед. 1999;159:473�479.
119. Новак Д, Касиелски М, Антцзак А, Пиетрас Т, Биаласиевицз П. Повећан садржај супстанци које реагују на тиобарбитурну киселину и водоник пероксида у кондензату издаха код пацијената са стабилном хроничном опструктивном болешћу плућа: нема значајног ефекта пушења цигарета. Респир Мед. 1999;93:389�396.
120. Келли ФЈ, Мудваи ИС. Оксидација протеина на интерфејсу ваздух-плућа. Амино киселине. 2003;25:375�396.
121. Деан РТ, Робертс ЦР, Јессуп В. Фрагментација екстрацелуларних и интрацелуларних полипептида слободним радикалима. Прог Цлин Биол Рес. 1985;180:341�350.
122. Кецк РГ. Употреба т-бутил хидропероксида као сонде за оксидацију метионина у протеинима. Анал Биоцхем. 1996;236:56�62.
123. Давиес КЈ. Оштећење и деградација протеина радикалима кисеоника. И. Општи аспекти. Ј Биол Цхем. 1987;262:9895�9901.
124. Стадтман ЕР. Оксидација протеина катализована металним јонима: биохемијски механизам и биолошке последице. Фрее Радиц Биол Мед.
1990;9:315�325.
125. Фуцци Л, Оливер ЦН, Цоон МЈ, Стадтман ЕР. Инактивација кључних метаболичких ензима реакцијама оксидације мешовите функције: могуће импликације у промету протеина и старењу. Проц Натл Ацад Сци УС А. 1983;80:1521�1525.
126. Стадтман ЕР, Московитз Ј, Левине РЛ. Оксидација метионинских остатака протеина: биолошке последице. Антиоксид Редок сигнал. 2003;5:577�582.
127. Стадтман ЕР, Левине РЛ. Оксидација слободних аминокиселина и аминокиселинских остатака у протеинима посредована слободним радикалима. Амино киселине. 2003;25:207�218.
128. Стадтман ЕР. Оксидација протеина код старења и болести повезаних са старењем. Анн НИ Ацад Сци. 2001;928:22�38.
129. Схацтер Е. Квантификација и значај оксидације протеина у биолошким узорцима. Друг Метаб Рев. 2000;32:307�326.
130. Поли Г, Леонардуззи Г, Биаси Ф, Цхиарпотто Е. Оксидативни стрес и сигнализација ћелија. Цурр Мед Цхем. 2004;11:1163�1182.
131. Неуфелд Г, Цохен Т, Генгриновитцх С, Полторак З. Фактор раста васкуларног ендотела (ВЕГФ) и његови рецептори. ФАСЕБ Ј. 1999;13:9.
132. Сундаресан М, Иу ЗКС, Ферранс ВЈ, Сулцинер ДЈ, Гуткинд ЈС, ет ал. Регулација стварања врста реактивног кисеоника у фибробластима помоћу Рац1. Биоцхем Ј. 1996;318:379-382.
133. Сун Т, Оберлеи ЛВ. Редок регулација транскрипционих активатора. Фрее Радиц Биол Мед. 1996;21:335�348.
134. Клатт П, Молина ЕП, Де Лацоба МГ, Падилла ЦА, Мартинез-Галестео Е, Барцена ЈА, Ламас С. Редок регулација ц-Јун ДНК везивања реверзибилном С-глутатиолацијом. ФАСЕБ Ј. 1999;13:1481-1490.
135. Реинаерт НЛ, Цклес К, Гуала АС, Воутерс ЕФ, ван дер Влиет А, Јанссен Хеинингер
ИМ. Ин ситу детекција С-глутатионилираних протеина након дериватизације цистеина катализоване глутаредоксином-1. Биоцхим Биопхис Ацта. 2006;1760:380�387.
136. Реинаерт НЛ, Воутерс ЕФ, Јанссен-Хеинингер ИМ. Модулација глутаредоксина-1
експресија на мишјем моделу алергијске болести дисајних путева. Ам Ј Респир Целл Мол Биол. 2007;36:147�151.
137. Филомени Г, Ротилио Г, Цириоло МР. Ћелијска сигнализација и редокс систем глутатиона. Биоцхем Пхармацол. 2002;64:1057�1064.
138. Панде В, Рамос МЈ. Молекуларно препознавање 15-деоксиделта (12,14) простагландина Ј(2) помоћу нуклеарног фактора-капа Б и других ћелијских протеина. Биоорг Мед Цхем Летт. 2005;15:4057�4063.
139. Перкинс НД. Интегрисање путева ћелијске сигнализације са НФ-каппаБ и ИКК функцијом. Нат Рев Мол Целл Биол. 2007;8:49�62.
140. Гилморе ТД. Увод у НФ-каппаБ: играчи, путеви, перспективе. Онкоген. 2006;25:6680�6684.
141. Хирота К, Мурата М, Сацхи И, Накамура Х, Такеуцхи Ј, Мори К, Иодои Ј. Различите улоге тиоредоксина у цитоплазми и у језгру. Двостепени механизам редокс регулације транскрипционог фактора НФ-каппаБ. Ј Биол Цхем. 1999;274:27891�27897.
142. Одељење ПА. Улога комплемента, хемокина и регулаторних цитокина у акутној повреди плућа. Анн НИ Ацад Сци. 1996;796:104�112.
143. Акира С, Кисхимото А. НФ-ИЛ6 и НФ-кБ у регулацији гена цитокина. Адв Иммунол. 1997;65:1�46.
144. Меиер М, Сцхрецк Р, Баеуерле ПА. Х2О2 и антиоксиданси имају супротне ефекте на активацију НФ-капа Б и АП-1 у нетакнутим ћелијама: АП-1 као секундарни фактор који реагује на антиоксиданс. ЕМБО Ј. 1993;12:2005–2015.
145. Абате Ц, Пател Л, Раусхер ФЈ, Цурран Т. Редок регулација фос и јун ДНК-везујуће активности ин витро. Наука. 1990;249:1157�1161.
146. Галтер Д, Михм С, Дроге В. Изразити ефекти глутатион дисулфида на факторе нуклеарне транскрипције кБ и протеин активатор-1. Еур Ј Биоцхем. 1994;221:639�648.
147. Хирота К, Матсуи М, Ивата С, Нисхииама А, Мори К, Иодои Ј. Транскрипциона активност АП-1 је регулисана директном асоцијацијом између тиоредоксина и Реф-1. Проц Натл Ацад Сци УС А. 1997;94: 3633�3638.
ИФМ-ов алат Пронађи практичара је највећа реферална мрежа у функционалној медицини, створена да помогне пацијентима да пронађу практичаре функционалне медицине било где у свету. ИФМ сертификовани практичари наведени су први у резултатима претраге, с обзиром на њихово опсежно образовање из функционалне медицине